
Precedenti studi di ricerca suggeriscono che l'L-aspartato, come l'L-glutammato, innesca l'attività eccitatoria sui neuroni. L-aspartato funziona con L-glutammato nelle vescicole sinaptiche delle sinapsi eccitatorie asimmetriche. Ma la concentrazione totale di questi nel cervello umano (0.96-1.62? Mol / grammo di peso umido), le loro concentrazioni extracellulari nella corteccia misurate mediante microdialisi (1.62? M per L-aspartato e 9.06? M per L-glutammato) e il loro apporto secondo l'immunoistochimica suggeriscono che l'L-aspartato è significativamente meno abbondante dell'L-glutammato. Inoltre, l'L-aspartato è un potente agonista per i recettori NMDA ma non per altri iGluR con un EC50 solo otto volte superiore a quello dell'L-glutammato. Gli EAAT che svolgono un ruolo fondamentale nell'assorbimento di tutto l'L-glutammato rilasciato vescicolare nel sistema nervoso centrale (SNC) richiedono anche l'utilizzo dell'L-aspartato. L'L-aspartato è forse meno essenziale dell'L-glutammato connesso all'attività eccitatoria totale associata agli iGluRs. Insieme al suo ruolo di neurotrasmettitore, come accennato in precedenza, l'L-aspartato è anche necessario come substrato per l'aspartato amino-transferasi che si trasforma in 2-ossoglutarato e L-glutammato per il trasporto alle vescicole corticali dei neuroni glutamatergici che possono anche conseguentemente e aumentare indirettamente il rilascio di L-glutammato.
Contenuti
Altre molecole nella segnalazione di glutammato
Una caratteristica che distingue i recettori NMDA dai diversi iGluR è che l'attivazione dei recettori NMDA richiede la connessione di un co-agonista alla regione di legame della glicina del recettore. Ad esempio, nella retina e nel midollo spinale, l'origine della glicina può fuoriuscire dalle sinapsi inibitorie glicinergiche. Ma, in diverse regioni del cervello con una maggiore espressione del recettore NMDA, come la formazione dell'ippocampo, mancano le reazioni associate ai recettori della glicina sensibili alla stricnina, almeno nei neuroni adulti, a dimostrazione dell'assenza di neurotrasmissioni inibitorie glicinergiche. Ma la glicina si trova nel fluido extracellulare dell'ippocampo a quantità basali di circa 1.5 μM, che è simile alla saturazione della regione di legame della glicina del recettore NMDA, sebbene questi possano essere regolati verso l'alto e verso il basso. L'origine della glicina extracellulare nell'ippocampo può essere neuroni che rilasciano glicina attraverso il trasportatore di amminoacidi alanina-serina-cisteina 1 (asc-1). Ma è stato anche dimostrato il rilascio di glicina da parte degli astrociti stimolato dalla depolarizzazione e dal kainato. Sono necessari ulteriori studi di ricerca per mostrare alla fine queste misure di esito.
Anche in precedenti studi di ricerca sul recettore NMDA e sulla sua co-attivazione da parte della glicina è emerso che gli amminoacidi D, in particolare la D-serina, sono potenti quasi quanto la glicina. Solo diversi anni dopo, è diventato ovvio che la D-serina si trova nel cervello di ratti e umani a circa un terzo della loro concentrazione di L-serina con una concentrazione assoluta di oltre 0.2 μmol / g di tessuto cerebrale. Utilizzando un antisiero per la D-serina, studi di ricerca hanno dimostrato che la D-serina dal cervello si trova solo negli astrociti e la sua fornitura si adatta all'espressione dei recettori NMDA. Inoltre, gli stessi ricercatori hanno dimostrato che la D-serina viene rilasciata dagli astrociti coltivati quando esposti a L-glutammato o kainato. L'abbondanza di D-serina è trovata dall'enzima degradante D-amminoacido ossidasi (DAO) che rivela una maggiore espressione nel hindbrain dove i livelli di D-serina sono ridotti così come l'enzima sintetico serina racemasi che crea D-serina da L- serina. La D-serina sembra essere immagazzinata nelle vescicole citoplasmatiche degli astrociti e può essere rilasciata per esocitosi. Il potenziamento a lungo termine dipende dal rilascio di D-serina dagli astrociti nelle fettine dell'ippocampo, suggerendo che questo amminoacido gioca sicuramente un ruolo fondamentale nella neurotrasmissione glutamatergica attraverso i recettori NMDA. Inoltre, nelle fettine dell'ippocampo, studi di ricerca hanno trovato, utilizzando D-serina e enzimi degradanti della glicina, che D-serina funziona come co-trasmettitore per i recettori NMDA sinaptici sui neuroni CA1, allo stesso modo la glicina funziona come co-agonista endogeno per i recettori NMDA extrasinaptici. I recettori NMDA sinaptici dei neuroni del giro dentato utilizzano la glicina anziché la D-serina come co-agonista.
Prese collettivamente, le misure di esito multistrato mostrano che l'L-aspartato non funziona semplicemente come un agonista sui recettori NMDA, ma anche la glicina e la D-serina svolgono un ruolo fondamentale nella neurotrasmissione glutamatergica nel cervello umano. Ma anche altre molecole hanno dimostrato di essere importanti modulatori della neurotrasmissione glutamatergica.
Glutammato attivato da altre molecole
L-omocisteato (L-HCA) ha somiglianze strutturali con L-glutammato. L'amminoacido non proteico è un prodotto di ossidazione dell'omocisteina che viene biosintetizzato dalla metionina nell'eliminazione del proprio gruppo metilico terminale ed è anche un intermedio della via di transolfurazione mediante la quale la metionina può essere convertita in cisteina attraverso la cistationina. I primi studi di ricerca hanno dimostrato che questo amminoacido può causare l'afflusso di calcio nei neuroni coltivati in modo sicuro ed efficace come l'L-glutammato. Inoltre, L-HCA ha rivelato un'aumentata affinità per i recettori NMDA rispetto ad altri iGluR nei saggi di legame associati alla sua capacità di causare eccitotossicità inibibile dall'antagonista del recettore NMDA e afflusso di sodio. Inoltre, L-HCA può attivare mGluR5 con la stessa efficienza dell'L-glutammato. L-HCA si trova nel cervello, tuttavia, è stato dimostrato che le concentrazioni sono circa 500 volte inferiori a quelle dell'L-glutammato e persino 100 volte inferiori rispetto a quelle dell'L-aspartato in diverse regioni del cervello del ratto. Durante la stimolazione indotta dal potassio, la scarica di L-HCA viene attivata dalle preparazioni di fette di cervello come dimostrato per L-aspartato e L-glutammato sebbene il rilascio assoluto di HCA sia circa 50 volte inferiore. Sorprendentemente, l'HCA è un inibitore competitivo molto efficiente della cistina e dell'assorbimento di L-glutammato attraverso il sistema antiportatore di cistina / glutammato x? C, l'attività che regola e gestisce le concentrazioni extracellulari di L-glutammato extrasinaptico nel cervello. Pertanto, l'impatto di L-HCA sull'attivazione di NMDA e di altri recettori di L-glutammato può anche fare affidamento sull'innesco di L-glutammato indotto da L-HCA attraverso il sistema x? C. L-HCA può svolgere un ruolo importante nella stimolazione complessiva dei recettori L-glutammato. Tuttavia, questo può cambiare enormemente in determinate condizioni, p. Es., Nei pazienti con terapia ad alte dosi con metotrexato, un farmaco antitumorale che, limitando la diidrofolato reduttasi, limita il riciclo catalizzato dal tetraidrofolato della metionina dall'omocisteina. Qui, concentrazioni di L-HCA superiori a 100 μM sono state dimostrate dal liquido cerebrospinale mentre L-HCA non era rilevabile nei soggetti di controllo. Sono ancora necessari ulteriori studi di ricerca per determinare queste misure di esito.
Ulteriori piccole molecole endogene che si ritiene influenzino la segnalazione di L-glutammato includono diversi intermedi del metabolismo del triptofano, come mostrato nella Figura 2. Attraverso l'attività dell'indoleamina 2,3-diossigenasi (IDO) o triptofano 2,3-diossigenasi (TDO), il triptofano viene trasformato in N-formil-L-chinurenina che viene successivamente trasformata in chinurenina (KYN) dalla formamidasi. Tre percorsi, due dei quali si collegano in una fase successiva, determinano un ulteriore metabolismo. Innanzitutto, attraverso l'attività della chinurenina aminotransferasi (KAT), il KYN viene convertito in acido chinurenico (KYNA). KYN può anche essere convertito in 3-idrossichinurenina (3HK) dalla chinurenina monoossigenasi (KMO), che può essere successivamente utilizzata come substrato dalla chinureninasi per la sintesi dell'acido 3-idrossiantranilico (3HANA). Inoltre, utilizzando KYN come substrato, la chinureninasi sviluppa acido antranilico (ANA), che mediante idrossilazione aspecifica può anche essere convertito in 3HANA. Secondo studi di ricerca, 3HANA funziona finalmente come substrato per la generazione di acido chinolinico (QUIN).
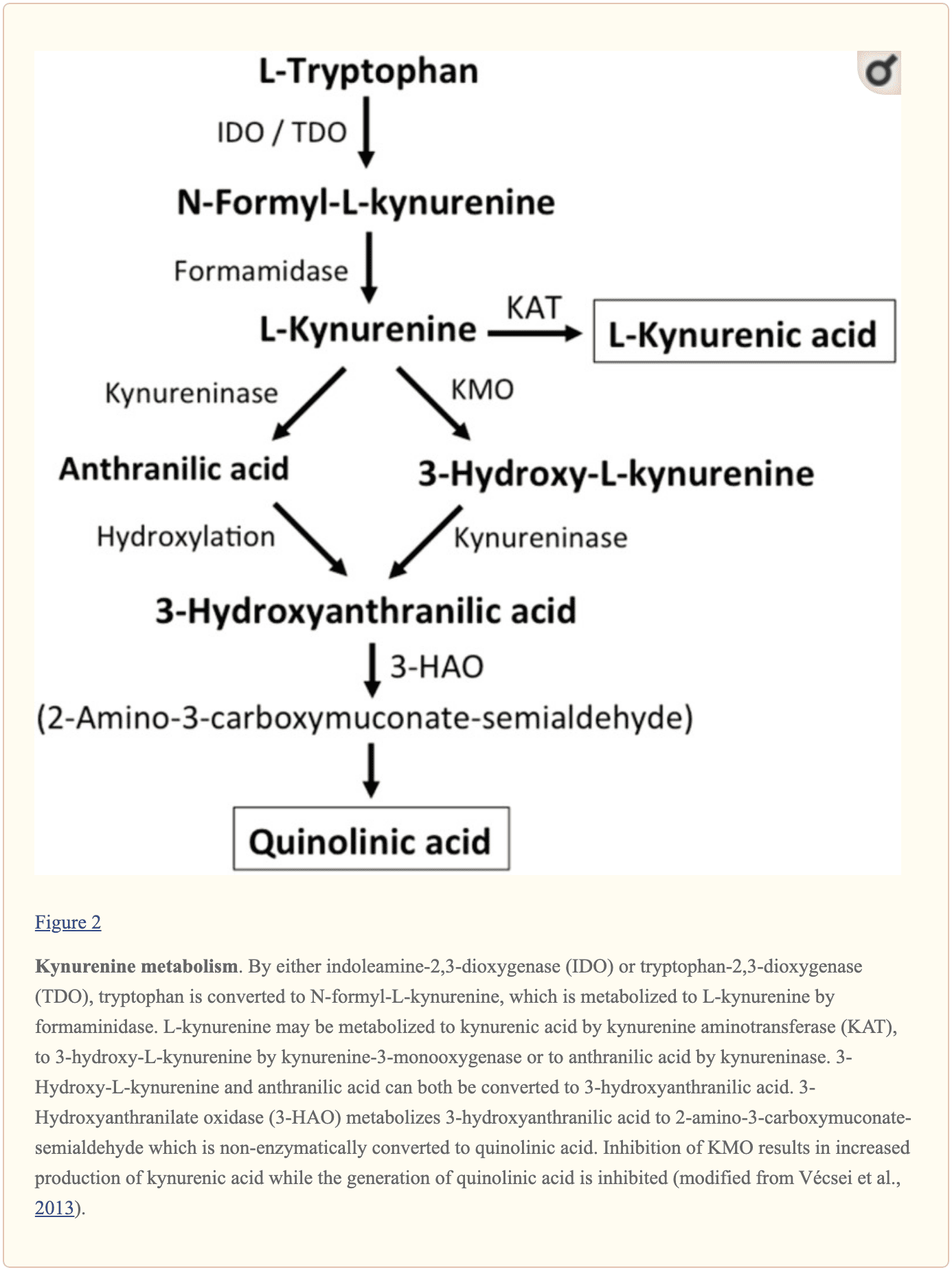
La concentrazione di triptofano nel cervello del ratto è di circa 25 nmol / g di peso umido e circa 400 volte inferiore all'L-glutammato e 100 volte inferiore all'L-aspartato. I livelli cerebrali dimostrati di chinurenine sono ancora più bassi con 0.4-1.6 nmol / g per QUIN, 0.01-0.07 nmol / ml per KYNA e 0.016 nmol / g per 3HANA. Circa il 40% del KYN cerebrale è sintetizzato localmente. I metaboliti del triptofano dimostrano un legame differenziale con le proteine plasmatiche e il loro trasporto attraverso la barriera che è molto diverso. KYN e 3HK vengono trasportati attraverso il grande sistema di trasporto di amminoacidi neutri L. Le kynurenine sembrano penetrare nel cervello umano per diffusione passiva. Inoltre, KYNA, 3HANA e soprattutto ANA si legano alle proteine del siero che alla fine ne restringono e limitano la diffusibilità attraverso la barriera ematoencefalica.
Studi di ricerca hanno dimostrato che QUIN, quando utilizzato ionoforeticamente nelle cellule di ratto, ha causato l'attivazione neuronale che è stata prevenuta da un antagonista del recettore NMDA, suggerendo che QUIN può funzionare come un agonista del recettore NMDA. Tuttavia, è stato dimostrato che l'EC50 per QUIN per attivare le correnti del recettore NMDA è circa 1000 volte superiore all'EC50 dell'L-glutammato. È stato dimostrato che l'iniezione intracerebrale di QUIN causa cambiamenti ultrastrutturali, neurochimici e comportamentali simili a quelli causati dagli agonisti del recettore NMDA. Il fatto che le concentrazioni di QUIN siano da circa 5000 a 15,000 volte inferiori alle concentrazioni di L-glutammato cerebrale rende improbabile che la modulazione della segnalazione del recettore NMDA da parte di QUIN svolga un ruolo essenziale. È stato dimostrato che KYNA funziona come un antagonista del recettore NMDA. Ma, sebbene l'infusione con l'inibitore di KMO Ro 61-8048 abbia migliorato di 10 volte le concentrazioni di KYNA extracellulare cerebrale, ciò non ha comportato un'inibizione della depolarizzazione neuronale mediata da NMDA, una scoperta che sfida la convinzione che KYNA a quantità quasi fisiologiche direttamente modula i recettori NMDA. In confronto, l'aumento di KYNA nel cervello indotto dall'inibitore KMO JM6 ha diminuito la concentrazione di L-glutammato cerebrale extracellulare. Inoltre, i livelli di KYNA dal liquido cerebrale extracellulare sono stati associati ai livelli di L-glutammato suggerendo che anche a livelli fisiologici o vicini a quelli fisiologici, KYNA modula il metabolismo dell'L-glutammato. Sia l'attivazione del recettore accoppiato alla proteina G GPR35 che l'inibizione dei recettori nicotinici dell'acetilcolina presinaptica? 7 sono suggerite nella riduzione indotta da KYNA del rilascio di L-glutammato. Per riassumere, sebbene QUIN e L-HCA siano presenti nel cervello umano, le loro concentrazioni discutono contro di loro con ruoli nella regolazione e nel mantenimento della neurotrasmissione. Al contrario, anche se i percorsi devono essere definiti in maggiore dettaglio, le prove supportano i livelli e l'opinione che la scarica possa essere modulata da KYNA e neurotrasmissione.

Il glutammato, insieme all'aspartato e ad altre molecole, sono alcuni dei principali neurotrasmettitori eccitatori nel cervello umano. Sebbene questi svolgano un ruolo fondamentale nella struttura generale e nella funzione del sistema nervoso centrale, compresi il cervello e il midollo spinale, quantità eccessive di altre molecole possono in definitiva innescare i recettori del glutammato. L'eccesso di glutammato può causare eccitotossicità che può portare a una varietà di problemi di salute, come il morbo di Alzheimer e altri tipi di malattie neurologiche. Il seguente articolo descrive come altre molecole possono attivare i recettori del glutammato. - Dr. Alex Jimenez DC, CCST Insight - Dr. Alex Jimenez DC, CCST Insight
Studi di ricerca suggeriscono che L-aspartato, come il L-glutammato, innesca l'attività eccitatoria. L-aspartato funziona con L-glutammato nelle vescicole sinaptiche delle sinapsi eccitatorie asimmetriche. Ma la concentrazione totale di questi nel cervello umano suggerisce che L-aspartato è significativamente meno abbondante di L-glutammato. Inoltre, L-aspartato è un potente agonista per i recettori NMDA ma non per altri iGluR con un EC50 appena otto volte superiore a quello del L-glutammato. Lo scopo delle nostre informazioni è limitato a problemi di chiropratica, salute muscoloscheletrica e nervosa, nonché articoli, argomenti e discussioni di medicina funzionale. Utilizziamo protocolli sanitari funzionali per il trattamento di lesioni o disturbi cronici del sistema muscolo-scheletrico. Per discutere ulteriormente l'argomento di cui sopra, non esitate a chiedere al Dr. Alex Jimenez o contattarci al numero 915-850-0900 .
A cura del Dr. Alex Jimenez
Riferimenti ï ¿½
- Lewerenz, Jan e Pamela Maher. Tossicità cronica del glutammato nelle malattie neurodegenerative: quali sono le prove? Frontiere in Neuroscienze, Frontiers Media SA, 16 dic. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4679930/.
Discussione argomento aggiuntiva: dolore cronico
Il dolore improvviso è una risposta naturale del sistema nervoso che aiuta a dimostrare possibili lesioni. Ad esempio, i segnali del dolore viaggiano da una regione lesa attraverso i nervi e il midollo spinale al cervello. Il dolore è generalmente meno grave poiché la lesione guarisce, tuttavia, il dolore cronico è diverso dal tipo medio di dolore. Con dolore cronico, il corpo umano continuerà a inviare segnali di dolore al cervello, indipendentemente dal fatto che la lesione sia guarita. Il dolore cronico può durare da alcune settimane a persino diversi anni. Il dolore cronico può influenzare enormemente la mobilità di un paziente e può ridurre la flessibilità, la forza e la resistenza.
Zoomer Plus neuronale per la malattia neurologica
 ï ¿½
ï ¿½
Il dottor Alex Jimenez utilizza una serie di test per aiutare a valutare le malattie neurologiche. Lo zoom neuraleTM Plus è una gamma di autoanticorpi neurologici che offre un riconoscimento specifico anticorpo-antigene. Il vibrante Neural ZoomerTM Plus è progettato per valutare la reattività di un individuo a 48 antigeni neurologici con connessioni a una varietà di malattie neurologicamente correlate. Il vibrante zoomer neuraleTM Plus mira a ridurre le condizioni neurologiche fornendo ai pazienti e ai medici una risorsa vitale per la diagnosi precoce del rischio e una maggiore attenzione alla prevenzione primaria personalizzata.
Formule per il supporto alla metilazione

XYMOGEN s Formule professionali esclusive sono disponibili attraverso selezionati professionisti sanitari autorizzati. La vendita su Internet e l'attualizzazione delle formule XYMOGEN sono severamente vietate
Orgogliosamente, Il dottor Alexander Jimenez rende le formule XYMOGEN disponibili solo per i pazienti sotto la nostra cura.
Si prega di chiamare il nostro ufficio in modo da poter assegnare una consulenza medica per l'accesso immediato.
Se sei un paziente di Injury Medical & Chiropractic Clinica, puoi chiedere informazioni su XYMOGEN chiamando 915-850-0900.
![]()
Per vostra comodità e revisione del XYMOGEN prodotti si prega di rivedere il seguente link. *XYMOGEN-Catalog-Scaricare ï ¿½
* Tutte le precedenti politiche XYMOGEN rimangono rigorosamente in vigore.
ï ¿½
Le informazioni qui riportate su "Neurologia funzionale: altre molecole nel glutammato" non intende sostituire un rapporto individuale con un professionista sanitario qualificato o un medico autorizzato e non è una consulenza medica. Ti incoraggiamo a prendere decisioni sanitarie basate sulla tua ricerca e collaborazione con un professionista sanitario qualificato.
Informazioni sul blog e discussioni sull'ambito
Il nostro ambito informativo è limitato a chiropratica, muscolo-scheletrico, medicine fisiche, benessere, contributo eziologico disturbi viscerosomatici all'interno di presentazioni cliniche, dinamiche cliniche associate ai riflessi somatoviscerali, complessi di sublussazione, problemi di salute sensibili e/o articoli, argomenti e discussioni di medicina funzionale.
Forniamo e presentiamo collaborazione clinica con specialisti di varie discipline. Ogni specialista è regolato dal proprio ambito di pratica professionale e dalla propria giurisdizione di licenza. Utilizziamo protocolli funzionali di salute e benessere per trattare e supportare la cura delle lesioni o dei disturbi del sistema muscolo-scheletrico.
I nostri video, post, argomenti, soggetti e approfondimenti trattano questioni cliniche, problemi e argomenti che riguardano e supportano direttamente o indirettamente il nostro ambito di pratica clinica.*
Il nostro ufficio ha ragionevolmente tentato di fornire citazioni di supporto e ha identificato lo studio o gli studi di ricerca pertinenti a sostegno dei nostri post. Forniamo copie degli studi di ricerca di supporto a disposizione degli organi di regolamentazione e del pubblico su richiesta.
Comprendiamo che copriamo questioni che richiedono una spiegazione aggiuntiva su come può essere d'aiuto in un particolare piano di assistenza o protocollo di trattamento; pertanto, per discutere ulteriormente l'argomento di cui sopra, non esitate a chiedere Dott. Alex Jimenez, DC, o contattaci al 915-850-0900.
Siamo qui per aiutare te e la tua famiglia.
Blessings
Il dottor Alex Jimenez DC MSACP, Marina Militare*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionalmedicine.com
Licenza come Dottore in Chiropratica (DC) in Texas & Nuovo Messico*
Licenza Texas DC n. TX5807, Licenza DC del New Mexico n. NM-DC2182
Autorizzato come infermiere registrato (RN*) in Florida
Licenza Florida Licenza RN # RN9617241 (controllo n. 3558029)
Stato compatto: Licenza multistato: Autorizzato ad esercitare in Stati 40*
Dott. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Il mio biglietto da visita digitale





