Supporto Clinico Neurofisiologico Back Clinic. El Paso, TX. Chiropratico, il Dott. Alexander Jimenez discute neurofisiologia clinica. Il dott. Jimenez esplorerà il significato clinico e le attività funzionali delle fibre nervose periferiche, del midollo spinale, del tronco cerebrale e del cervello nel contesto dei disturbi viscerali e muscolo-scheletrici. I pazienti acquisiranno una conoscenza avanzata dell'anatomia, della genetica, della biochimica e della fisiologia del dolore in relazione a varie sindromi cliniche. Verranno incorporati biochimica nutrizionale correlata alla nocicezione e al dolore. E l'applicazione di queste informazioni nei programmi di terapia sarà enfatizzata.
Il nostro team è molto orgoglioso di offrire alle nostre famiglie e ai pazienti feriti solo protocolli di trattamento comprovati. Insegnando il benessere olistico completo come stile di vita, cambiamo anche non solo la vita dei nostri pazienti, ma anche le loro famiglie. Lo facciamo in modo da poter raggiungere tutti gli El Pasoani che hanno bisogno di noi, indipendentemente dai problemi di accessibilità. Per le risposte a qualsiasi domanda tu possa avere, chiama il Dr. Jimenez al 915-850-0900.
"Regole di decisione clinica, classificazione del dolore spinale e previsione dell'outcome del trattamento: una discussione sulle relazioni recenti nella letteratura riabilitativa"
Astratto
Le regole di decisione clinica sono una presenza sempre più comune nella letteratura biomedica e rappresentano una strategia per migliorare il processo decisionale clinico per migliorare l'efficienza e l'efficacia dell'assistenza sanitaria. Nel contesto della ricerca riabilitativa, le regole di decisione clinica sono state prevalentemente finalizzate a classificare i pazienti prevedendo la loro risposta al trattamento a terapie specifiche. Tradizionalmente, le raccomandazioni per lo sviluppo di regole di decisione clinica propongono un processo a più fasi (derivazione, convalida, analisi dell'impatto) utilizzando la metodologia definita. Gli sforzi di ricerca volti a sviluppare una regola di decisione clinica basata sulla diagnosi si sono allontanati da questa convenzione. Pubblicazioni recenti in questa linea di ricerca hanno utilizzato la guida alla decisione clinica basata sulla diagnosi terminologica modificata. Le modifiche alla terminologia e alla metodologia che circondano le regole di decisione clinica possono rendere più difficile per i medici riconoscere il livello di evidenza associato a una regola di decisione e capire come questa evidenza dovrebbe essere implementata per informare l'assistenza al paziente. Forniamo una breve panoramica dello sviluppo delle regole di decisione clinica nel contesto della letteratura riabilitativa e due articoli specifici recentemente pubblicati in Chiropratica e Terapie manuali.
Regole predizione clinica
L'assistenza sanitaria ha subito un importante cambiamento di paradigma verso la pratica basata sull'evidenza. Un approccio pensato per migliorare il processo decisionale clinico integrando le migliori evidenze disponibili con l'esperienza clinica e le preferenze dei pazienti.
In definitiva, l'obiettivo della pratica basata sull'evidenza è migliorare l'assistenza sanitaria. Tuttavia, la traduzione delle prove scientifiche in pratica si è rivelata un'impresa impegnativa.
Le regole di decisione clinica (CDR), note anche come regole di previsione clinica, sono sempre più comuni nella letteratura riabilitativa.
Questi sono strumenti progettati per informare il processo decisionale clinico identificando potenziali predittori dell'esito del test diagnostico, della prognosi o della risposta terapeutica.
Nella letteratura riabilitativa, le CDR sono più comunemente utilizzate per prevedere la risposta di un paziente al trattamento. Sono stati proposti per identificare sottogruppi clinicamente rilevanti di pazienti che presentano disturbi altrimenti eterogenei come collo non specifico o basso mal di schiena, che è la prospettiva su cui intendiamo concentrarci.
Regole predizione clinica
La capacità di classificare o sottoraggruppare i pazienti con disturbi eterogenei come il dolore spinale è stata evidenziata come una priorità della ricerca e, di conseguenza, il fulcro di molti sforzi di ricerca. L'attrattiva di tali approcci di classificazione è il loro potenziale per una migliore efficienza ed efficacia del trattamento abbinando i pazienti a terapie ottimali. In passato, la classificazione dei pazienti si è basata su approcci impliciti fondati nella tradizione o osservazioni non sistematiche. L'uso dei CDR per informare la classificazione è un tentativo di un approccio più basato sull'evidenza, meno dipendente da teorie infondate.
CDR sono sviluppati in un processo a più fasi che coinvolge studi di derivazione, convalida e analisi di impatto, ciascuno con uno scopo definito e criteri metodologici. Come per tutte le forme di evidenza utilizzate per prendere decisioni sui pazienti, l'attenzione alla metodologia di studio appropriata è fondamentale per valutare i potenziali benefici dell'implementazione.
Benefici delle regole di previsione clinica
Può ospitare più fattori di quanti il cervello umano possa prendere in considerazione
Il modello CDR / CPR darà sempre lo stesso risultato (equazione matematica)
Può essere più accurato del giudizio clinico.
Usi clinici delle regole di previsione clinica
Diagnosi Probabilità di pretest
Prognosi Prevedere il rischio di esiti della malattia
In definitiva, l'utilità di un CDR non risiede nella sua accuratezza, ma nella sua capacità di migliorare i risultati clinici e aumentare l'efficienza delle cure.[15] Anche quando un CDR dimostra un'ampia convalida, ciò non garantisce che cambierà il processo decisionale clinico o che i cambiamenti che produce si tradurranno in una migliore assistenza.
I cambiamenti che produce si tradurranno in una migliore cura. McGinn et al.[2] individuato tre spiegazioni per il fallimento di un CDR in questa fase. In primo luogo, se il giudizio del medico è accurato quanto una decisione informata dal CDR, non vi è alcun vantaggio dal suo utilizzo. In secondo luogo, l'applicazione di un CDR può comportare calcoli o procedure ingombranti che scoraggiano i medici dall'utilizzare il CDR. Terzo, l'utilizzo del CDR potrebbe non essere fattibile in tutti gli ambienti o circostanze. Inoltre, includiamo la realtà che gli studi sperimentali possono coinvolgere pazienti che non sono del tutto rappresentativi di quelli osservati nelle cure di routine e che ciò può limitare il valore effettivo di una CDR. Pertanto, per comprendere appieno l'utilità di un CDR e la sua capacità di migliorare l'assistenza sanitaria, è necessario intraprendere un esame pragmatico della sua fattibilità e impatto quando applicato in un ambiente che riflette la pratica del mondo reale. Questo può essere intrapreso con diversi disegni di studio come studi randomizzati, studi randomizzati a cluster o altri approcci come l'esame dell'impatto di un CDR prima e dopo la sua implementazione.
Prevalenza dei metodi di classificazione per i pazienti con compromissione lombare utilizzando le sindromi di McKenzie, il pattern di dolore, la manipolazione e le regole di previsione clinica di stabilizzazione.
Gli obiettivi erano (1) per determinare la proporzione di pazienti con menomazioni lombari che potevano essere classificati all'assunzione dalle sindromi McKenzie (McK) e dalla classificazione del pattern del dolore (PPC) utilizzando metodi di valutazione e manipolazione meccanica e di diagnosi meccanica (MDT) e di stabilizzazione regole (CPR) e (2) per ciascuna categoria CPR uomo o CPR Stab, determinare i tassi di prevalenza di classificazione utilizzando McK e PPC.
Le RCP sono sofisticati modelli probabilistici e prognostici in cui un gruppo di caratteristiche del paziente identificate e segni e sintomi clinici sono statisticamente associati a una previsione significativa degli esiti dei pazienti.
I ricercatori hanno sviluppato due RCP separate per identificare i pazienti che avrebbero risposto favorevolmente alla manipolazione.33,34 Flynn et al. sviluppato la RCP di manipolazione originale utilizzando cinque criteri, ovvero nessun sintomo al di sotto del ginocchio, recente insorgenza di sintomi (<16 giorni), punteggio basso del questionario sulla convinzione di evitare la paura36 per il lavoro (<19), ipomobilità della colonna lombare e interno dell'anca rotazione ROM (>35 per almeno un'anca).33
La RCP di Flynn è stata successivamente modificata da Fritz et al. a due criteri, che includevano l'assenza di sintomi al di sotto del ginocchio e l'insorgenza recente dei sintomi (<16 giorni), come alternativa pragmatica per ridurre l'onere clinico per identificare i pazienti nelle cure primarie che hanno maggiori probabilità di rispondere alla manipolazione della spinta.34 positivamente
"Potentia.l Insidie delle regole di previsione clinica"
Quali sono le regole di previsione clinica?
Una regola di previsione clinica (CPR) è una combinazione di risultati clinici che hanno dimostrato statisticamente una prevedibilità significativa nel determinare una condizione selezionata o la prognosi di un paziente a cui è stato fornito un trattamento specifico 1,2. Le RCP sono create utilizzando metodi statistici multivariati, sono progettate per esaminare la capacità predittiva di raggruppamenti selezionati di variabili cliniche3,4 e hanno lo scopo di aiutare i medici a prendere decisioni rapide che normalmente possono essere soggette a pregiudizi sottostanti5. Le regole sono di natura algoritmica e coinvolgono informazioni condensate che identificano il minor numero di indicatori statisticamente diagnostici alla condizione mirata6.
Le regole di previsione clinica sono generalmente sviluppate utilizzando un metodo in 3 fasi14. In primo luogo, le RCP ci hanno derivato in modo prospettico-
metodi statistici multivariati per esaminare la capacità predittiva di raggruppamenti selezionati di variabili cliniche3. La seconda fase prevede la convalida della RCP in uno studio controllato randomizzato per ridurre il rischio che i fattori predittivi sviluppati durante la fase di derivazione siano selezionati per caso14. La terza fase prevede la conduzione di un'analisi dell'impatto per determinare in che modo la RCP migliora l'assistenza, riduce i costi e definisce con precisione l'obiettivo prefissato14.
Sebbene sia poco dibattuto che le RCP accuratamente costruite possano migliorare la pratica clinica, per quanto ne so, non ci sono linee guida che specificano i requisiti metodologici per i RCP per infusione in tutti gli ambienti di pratica clinica. Sono state create linee guida per migliorare il rigore della progettazione dello studio e del reporting. Il seguente editoriale delinea le potenziali insidie metodologiche nei CPR che potrebbero indebolire significativamente la trasferibilità dell'algoritmo. Nell'ambito della riabilitazione, la maggior parte delle RCP sono state prescrittive; quindi, i miei commenti qui riflettono le RCP prescrittive.
Insidie metodologiche
Le RCP sono progettate per specificare un insieme omogeneo di caratteristiche da una popolazione eterogenea di pazienti consecutivi selezionati in modo prospettico5,15. In genere, la popolazione applicabile risultante è un piccolo sottoinsieme di un campione più ampio e può rappresentare solo una piccola percentuale dell'effettivo carico di lavoro giornaliero del medico. L'impostazione e la posizione del campione più ampio dovrebbero essere generalizzabili15,16 e successivi studi di validità richiedono la valutazione della RCP in diversi gruppi di pazienti, in ambienti diversi e con un tipico gruppo di pazienti visto dalla maggior parte dei medici16. Poiché molte RCP sono sviluppate sulla base di un gruppo molto distinto che può o meno riflettere una popolazione tipica di pazienti, la trasportabilità dello spettro17 di molti algoritmi RCP attuali può essere limitata.
Le regole di previsione clinica utilizzano misure di esito per determinare l'efficacia dell'intervento. Le misure di esito devono avere un'unica definizione operativa5 e richiedere una reattività sufficiente per catturare realmente il cambiamento appropriato nella condizione14; inoltre, queste misure dovrebbero avere un cut-off score16,18 ben costruito ed essere raccolte da un amministratore in cieco15. La selezione di un punteggio di ancoraggio appropriato per la misurazione del cambiamento effettivo è attualmente dibattuta19-20. La maggior parte delle misure di esito utilizza un questionario basato sul ricordo del paziente come un punteggio globale di cambiamento del punteggio (GRoC), che è appropriato se usato a breve termine, ma soffre di bias di richiamo quando usato in analisi a lungo termine19-21.
Un potenziale svantaggio per le RCP è l'incapacità di mantenere la qualità dei test e delle misure utilizzate come predittori nell'algoritmo. Pertanto, il test prospettico e le misure dovrebbero essere indipendenti l'uno dall'altro durante la modellazione16; ciascuno dovrebbe essere eseguito in modo significativo e accettabile4; i medici o gli amministratori dei dati dovrebbero essere all'oscuro delle misure e delle condizioni degli esiti del paziente22.
fonti
Potenziali insidie delle regole di previsione clinica; The Journal of Manual & Manipulative Therapy Volume 16 Numero Due [69]
Jeffrey J Hebert e Julie M Fritz; Regole di decisione clinica, classificazione del dolore spinale e previsione dell'outcome del trattamento: una discussione sulle relazioni recenti nella letteratura di riabilitazione
La depressione è uno dei problemi di salute mentale più comuni negli Stati Uniti. La ricerca attuale suggerisce che la depressione deriva da una combinazione di aspetti genetici, biologici, ecologici e psicologici. La depressione è uno dei principali disordini psichiatrici in tutto il mondo con una significativa tensione economica e psicologica sulla società. Fortunatamente, la depressione, anche i casi più gravi, possono essere trattati. Quanto prima può iniziare il trattamento, tanto più efficace è.
Di conseguenza, tuttavia, vi è la necessità di biomarcatori robusti che contribuiranno a migliorare la diagnosi al fine di accelerare il processo di scoperta di farmaci e / o farmaci per ciascun paziente affetto dal disturbo. Questi sono indicatori fisiologici oggettivi, periferici, la cui presenza può essere utilizzata per prevedere la probabilità di insorgenza o esistenza di depressione, stratificarsi secondo gravità o sintomatologia, indicare predire e prognosi o monitorare la risposta agli interventi terapeutici. Lo scopo del seguente articolo è di dimostrare le intuizioni recenti, le sfide attuali e le prospettive future relative alla scoperta di una varietà di biomarcatori per la depressione e come questi possono aiutare a migliorare la diagnosi e il trattamento.
Biomarcatori per la depressione: approfondimenti recenti, sfide attuali e prospettive future
Astratto
Una pletora di ricerche ha implicato centinaia di putativi biomarcatori per la depressione, ma non ha ancora chiarito completamente i loro ruoli nella malattia depressiva o stabilito ciò che è anormale in cui i pazienti e in che modo le informazioni biologiche possono essere utilizzate per migliorare diagnosi, trattamento e prognosi. Questa mancanza di progresso è parzialmente dovuta alla natura e all'eterogeneità della depressione, in combinazione con l'eterogeneità metodologica all'interno della letteratura di ricerca e la vasta gamma di biomarcatori con potenziale, la cui espressione varia spesso in base a molti fattori. Esaminiamo la letteratura disponibile, che indica che i marcatori coinvolti nei processi infiammatori, neurotrofici e metabolici, così come i componenti del sistema neurotrasmettitore e neuroendocrino, rappresentano candidati molto promettenti. Questi possono essere misurati attraverso valutazioni genetiche ed epigenetiche, trascrittomiche e proteomiche, metabolomiche e di neuroimaging. L'uso di nuovi approcci e programmi di ricerca sistematici è ora necessario per determinare se e quali biomarcatori possono essere usati per predire la risposta al trattamento, stratificare i pazienti in trattamenti specifici e sviluppare obiettivi per nuovi interventi. Concludiamo che ci sono molte promesse per ridurre il peso della depressione attraverso l'ulteriore sviluppo e l'espansione di questi percorsi di ricerca.
parole chiave:disturbo dell'umore, disturbo depressivo maggiore, infiammazione, risposta al trattamento, stratificazione, medicina personalizzata
Introduzione
Sfide nei disturbi della salute mentale e dell'umore
Anche se la psichiatria ha un onere correlato alla malattia più grande di qualsiasi altra categoria di diagnostica medica, 1 è ancora una disparità di stima tra salute fisica e mentale in molti campi, tra cui finanziamento della ricerca2 e pubblicazione.3 Tra le difficoltà che la salute mentale affronta è una mancanza di consenso che circonda la classificazione, la diagnosi e il trattamento che deriva da una comprensione incompleta dei processi alla base di questi disturbi. Questo è molto evidente nei disturbi dell'umore, la categoria che comprende il singolo più grande onere nella salute mentale.3 Il disturbo dell'umore più diffuso, il disturbo depressivo maggiore (MDD), è una malattia complessa ed eterogenea in cui può verificarsi fino a 60% di pazienti un certo grado di resistenza al trattamento che prolunga e peggiora gli episodi.4 Per i disturbi dell'umore, e nel più ampio campo della salute mentale, i risultati del trattamento verrebbero probabilmente migliorati dalla scoperta di sottotipi robusti e omogenei all'interno di (e attraverso) categorie diagnostiche, mediante i quali i trattamenti potrebbe essere stratificato. In riconoscimento di ciò, sono ora in corso iniziative globali per delineare sottotipi funzionali, come i criteri del dominio di ricerca. 5 È stato ipotizzato che i marcatori biologici siano candidati prioritari per i disturbi mentali sottotipizzati. 6
Migliorare la risposta ai trattamenti per la depressione
Nonostante una vasta gamma di opzioni di trattamento per la depressione maggiore, solo circa un terzo dei pazienti con MDD ottiene la remissione anche quando riceve un trattamento antidepressivo ottimale secondo le linee guida di consenso e l'utilizzo di cure basate sulla misurazione, e le percentuali di risposta al trattamento sembrano diminuire con ogni nuovo trattamento .7 Inoltre, la depressione resistente al trattamento (TRD) è associata ad un aumento della compromissione funzionale, della mortalità, della morbilità e degli episodi ricorrenti o cronici a lungo termine. 8,9 Pertanto, ottenere miglioramenti nella risposta al trattamento in qualsiasi fase clinica offrirebbe maggiori benefici per i risultati complessivi in depressione. Nonostante il notevole onere imputabile alla TRD, la ricerca in questo settore è stata scarsa. Le definizioni di TRD non sono standardizzate, nonostante i precedenti tentativi: 4 alcuni criteri richiedono solo una prova di trattamento che non riesce a ottenere una riduzione del punteggio dei sintomi 50% (da una misura convalidata di gravità della depressione), mentre altri richiedono il mancato raggiungimento della remissione completa o mancata risposta ad almeno due antidepressivi adeguatamente testati di diverse classi all'interno di un episodio da considerare TRD.4,10 Inoltre, la stadiazione e la previsione della resistenza del trattamento è migliorata aggiungendo le caratteristiche cliniche chiave di gravità e cronicità al numero di trattamenti falliti. 9,11 Tuttavia, questa incoerenza nella definizione rende interpretare la letteratura di ricerca su TRD un compito ancora più complesso.
Al fine di migliorare la risposta ai trattamenti, è chiaramente utile identificare i fattori di rischio predittivi di non risposta. Sono stati caratterizzati alcuni predittori generali di TRD, tra cui la mancanza di completa remissione dopo episodi precedenti, ansia comorbida, suicidalità e insorgenza precoce della depressione, nonché personalità (particolarmente bassa estroversione, bassa gratificazione e alto nevroticismo) e fattori genetici.12 Questi risultati sono corroborati da revisioni che sintetizzano le evidenze separatamente per il trattamento farmacologico 13 e psicologico 14 per la depressione. Gli antidepressivi e le terapie cognitivo-comportamentali mostrano un'efficacia circa comparabile, 15, ma a causa dei loro diversi meccanismi di azione si potrebbe prevedere che abbiano diversi predittori di risposta. Mentre i traumi della prima infanzia sono stati a lungo associati a esiti clinici più poveri e ridotte risposte al trattamento, le prime indicazioni di 16 suggeriscono che le persone con una storia di trauma infantile potrebbero rispondere meglio alle terapie psicologiche che farmacologiche. 17 Nonostante ciò, prevale l'incertezza e poca personalizzazione o la stratificazione del trattamento ha raggiunto la pratica clinica.18
Questa recensione si concentra sulle prove a sostegno dell'utilità dei biomarcatori come strumenti clinici potenzialmente utili per migliorare la risposta al trattamento per la depressione.
Biomarcatori: sistemi e fonti
I biomarcatori forniscono un potenziale bersaglio per identificare i fattori predittivi di risposta a vari interventi.19 Le prove fino ad oggi suggeriscono che i marcatori che riflettono l'attività di sistemi infiammatori, neurotrasmettitori, neurotrofici, neuroendocrini e metabolici possono essere in grado di prevedere gli esiti di salute mentale e fisica negli individui attualmente depressi ma c'è molta incongruenza tra i risultati. 20 In questa recensione, ci concentriamo su questi cinque sistemi biologici.
Per ottenere una piena comprensione dei percorsi molecolari e del loro contributo nei disturbi psichiatrici, è ora considerato importante valutare più `` livelli '' biologici, in quello che viene comunemente definito un approccio `` omico ''. La Figura 21 fornisce una rappresentazione dei diversi livelli biologici a cui ciascuno dei cinque sistemi può essere valutato e le potenziali fonti di marcatori su cui queste valutazioni possono essere intraprese. Tuttavia, si noti che mentre ogni sistema può essere ispezionato a ciascun livello omico, le fonti di misurazione ottimali variano chiaramente a ciascun livello. Ad esempio, la neuroimaging fornisce una piattaforma per la valutazione indiretta della struttura o della funzione cerebrale, mentre gli esami delle proteine nel sangue valutano direttamente i marcatori. La trascrittomica1 e la metabolomica22 sono sempre più popolari, offrendo la valutazione di un numero potenzialmente enorme di marcatori, e il progetto Human Microbiome sta ora tentando di identificare tutti i microrganismi e la loro composizione genetica all'interno degli esseri umani.23 Nuove tecnologie stanno migliorando la nostra capacità di misurarli, anche attraverso fonti aggiuntive ; per esempio, gli ormoni come il cortisolo possono ora essere dosati sui capelli o sulle unghie (fornendo un'indicazione cronica) o sul sudore (fornendo una misurazione continua), 24 così come nel sangue, nel liquido cerebrospinale, nelle urine e nella saliva.
Dato il numero di presunte fonti, livelli e sistemi coinvolti nella depressione, non sorprende che la scala dei biomarcatori con potenziale traslazionale sia ampia. In particolare, quando si considerano le interazioni tra i marcatori, è forse improbabile che l'esame di singoli biomarcatori in isolamento produca risultati fruttuosi per il miglioramento della pratica clinica. Schmidt et al.26 hanno proposto l'uso di pannelli di biomarcatori e, successivamente, Brand et al27 hanno delineato una bozza di panel basata su precedenti evidenze cliniche e precliniche per MDD, identificando 16 bersagli di biomarcatori `` forti '', ognuno dei quali raramente è un singolo marcatore. Comprendono un volume ridotto della materia grigia (nelle regioni dell'ippocampo, della corteccia prefrontale e dei gangli della base), alterazioni del ciclo circadiano, ipercortisolismo e altre rappresentazioni di iperattivazione dell'asse ipotalamo-ipofisario-surrenale (HPA), disfunzione tiroidea, riduzione della dopamina, noradrenalina o acido 5-idrossindolacetico , aumento del glutammato, aumento della superossido dismutasi e della perossidazione lipidica, adenosina ciclica attenuata 3?, 5? -monofosfato e attività della proteina chinasi attivata da mitogeno, aumento delle citochine proinfiammatorie, alterazioni del triptofano, chinurenina, insulina e polimorfismi genetici specifici. Questi indicatori non sono stati concordati per consenso e potrebbero essere misurati in vari modi; è chiaro che un lavoro mirato e sistematico deve affrontare questo enorme compito al fine di dimostrare i loro benefici clinici.
Obiettivi di questa recensione
Come una revisione volutamente ampia, questo articolo cerca di determinare le esigenze complessive della ricerca sui biomarcatori nella depressione e la misura in cui i biomarcatori hanno un reale potenziale di traduzione per migliorare la risposta ai trattamenti. Iniziamo discutendo i risultati più importanti ed entusiasmanti in questo campo e indirizziamo il lettore a recensioni più specifiche relative a marcatori e confronti pertinenti. Delineiamo le attuali sfide affrontate alla luce delle prove, in combinazione con le esigenze di ridurre il peso della depressione. Infine, guardiamo avanti agli importanti percorsi di ricerca per affrontare le sfide attuali e le loro implicazioni per la pratica clinica.
Approfondimenti recenti
La ricerca di biomarcatori clinicamente utili per le persone con depressione ha generato ampie indagini nell'ultimo mezzo secolo. I trattamenti più comunemente usati sono stati concepiti dalla teoria monoamine della depressione; successivamente, le ipotesi neuroendocrine guadagnarono molta attenzione. In anni più recenti, la ricerca più prolifica ha circondato l'ipotesi infiammatoria della depressione. Tuttavia, un gran numero di articoli di revisione pertinenti si sono concentrati su tutti e cinque i sistemi; consulta la Tabella 1 e sotto per una raccolta di informazioni recenti sui sistemi di biomarker. Sebbene siano state misurate a molti livelli, le proteine derivate dal sangue sono state esaminate più ampiamente e forniscono una fonte di biomarker che è conveniente, economica e può essere più vicina al potenziale di traslazione rispetto ad altre fonti; quindi, maggiori dettagli vengono dati ai biomarcatori che circolano nel sangue.
In una recente revisione sistematica, Jani et al.20 hanno esaminato i biomarcatori del sangue periferico per la depressione in associazione con i risultati del trattamento. Dei soli 14 studi inclusi (cercati fino all'inizio del 2013), sono stati studiati 36 biomarcatori di cui 12 erano predittori significativi di indici di risposta mentale o fisica in almeno un'indagine. Quelli identificati come potenziali fattori di rischio per la mancata risposta includevano proteine infiammatorie: bassa interleuchina (IL) -12p70, rapporto tra conta dei linfociti e dei monociti; marker neuroendocrini (desametasone non soppressione del cortisolo, cortisolo alto circolante, ridotto ormone stimolante la tiroide); marker dei neurotrasmettitori (bassi livelli di serotonina e noradrenalina); fattori metabolici (colesterolo lipoproteico a bassa densità ad alta densità) e neurotrofici (proteina B legante il calcio S100 ridotta). Oltre a ciò, altre revisioni hanno riportato associazioni tra biomarcatori aggiuntivi e risultati del trattamento.19,28-30 Una breve descrizione dei marcatori putativi in ciascun sistema è delineata nelle sezioni successive e nella Tabella 2.
Risultati infiammatori in depressione
Dal seminale articolo di Smith che delinea l'ipotesi dei macrofagi31, questa letteratura consolidata ha trovato livelli aumentati di vari marcatori proinfiammatori in pazienti depressi, che sono stati ampiamente rivisti.32 Dodici proteine infiammatorie sono state valutate in meta-analisi che confrontano depressi e sani popolazioni di controllo. 37 38
IL-6 (P <0.001 in tutte le meta-analisi; 31 studi inclusi) e CRP (P <0.001; 20 studi) appaiono frequentemente e in modo affidabile elevati nella depressione.40 Un elevato fattore di necrosi tumorale alfa (TNF?) È stato identificato nei primi studi (P <0.001), 38 ma una sostanziale eterogeneità ha reso questo inconcludente quando si tiene conto di indagini più recenti (31 studi) .40 IL-1? è ancora più inconcludentemente associato alla depressione, con meta-analisi che suggeriscono livelli più elevati di depressione (P = 0.03), 41 livelli elevati solo negli studi europei42 o nessuna differenza rispetto ai controlli.40 Nonostante ciò, un recente articolo ha suggerito particolari implicazioni traslazionali per IL- 1?, 44 supportato da un effetto estremamente significativo di elevata IL-1? acido ribonucleico che predice una scarsa risposta agli antidepressivi; 45 altri risultati sopra riguardano le citochine ematiche circolanti. La chemochina monocita chemoattractant protein-1 ha mostrato aumenti nei partecipanti depressi in una meta-analisi.39 Le interleuchine IL-2, IL-4, IL-8, IL-10 e l'interferone gamma non erano significativamente differenti tra pazienti depressi e controlli a livello meta-analitico, ma hanno comunque dimostrato un potenziale in termini di alterazione con il trattamento: IL-8 è stato segnalato come elevato in quelli con depressione grave prospetticamente e trasversalmente, 46 diversi modelli di cambiamento di IL-10 e interferone gamma durante il trattamento si sono verificati tra pazienti che hanno risposto precocemente rispetto a quelli che non hanno risposto, 47 mentre IL-4 e IL-2 sono diminuiti in linea con la remissione dei sintomi.48 Nelle meta-analisi, sono state dimostrate piccole diminuzioni insieme al trattamento per IL-6, IL-1 ?, IL- 10 e CRP.43,49,50 Inoltre, TNF? può ridursi solo con il trattamento nei responder e un indice di marker composito può indicare un aumento dell'infiammazione nei pazienti che successivamente non rispondono al trattamento.43 È degno di nota, tuttavia, che quasi tutta la ricerca che esamina le proteine infiammatorie e la risposta al trattamento utilizza studi di trattamento farmacologico . Pertanto, almeno alcune alterazioni infiammatorie durante il trattamento sono probabilmente attribuibili agli antidepressivi. Gli effetti infiammatori precisi di diversi antidepressivi non sono stati ancora stabiliti, ma l'evidenza che utilizza i livelli di PCR suggerisce che gli individui rispondono in modo diverso a trattamenti specifici basati sull'infiammazione di base: Harley et al51 hanno riportato una CRP pre-trattamento elevata che predice una scarsa risposta alla terapia psicologica (cognitiva, comportamentale o interpersonale). psicoterapia), ma una buona risposta alla nortriptilina o alla fluoxetina; Uher et al52 hanno replicato questo risultato per la nortriptilina e hanno identificato l'effetto opposto per escitalopram. Al contrario, Chang et al53 hanno riscontrato una PCR più alta nei primi pazienti responsivi alla fluoxetina o alla venlafaxina rispetto ai non responder. Inoltre, i pazienti con TRD e CRP alta hanno risposto meglio al TNF? antagonista infliximab rispetto a quelli con livelli nel range di normalità. 54
Insieme, l'evidenza suggerisce che anche quando si controllano fattori come l'indice di massa corporea (BMI) e l'età, le risposte infiammatorie appaiono aberranti in circa un terzo dei pazienti con depressione.55,56 Il sistema infiammatorio, tuttavia, è estremamente complesso, e ci sono numerosi biomarcatori che rappresentano diversi aspetti di questo sistema. Recentemente, nuove citochine e chemochine hanno dato evidenza di anormalità nella depressione. Questi includono: proteina inibitoria dei macrofagi 1a, IL-1a, IL-7, IL-12p70, IL-13, IL-15, eotassina, fattore stimolante le colonie dei macrofagi dei granulociti, 57 IL-5,58 IL-16,59 IL-17,60 proteina chemiotattica dei monociti -4,61 timo e chemochina regolata per attivazione, 62 eotaxin-3, TNFb, 63 interferone gamma-indotta proteina 10,64 siero amiloide A, 65 solubile molecola di adesione intracellulare66 e molecola di adesione cellulare vascolare solubile 1.67
Risultati del fattore di crescita in depressione
Alla luce della potenziale importanza dei fattori di crescita non neurotrofici (come quelli relativi all'angiogenesi), ci riferiamo ai biomarcatori neurogenici sotto la più ampia definizione di fattori di crescita.
Il fattore neurotrofico derivato dal cervello (BDNF) è il più frequentemente studiato di questi. Molteplici meta-analisi dimostrano attenuazioni della proteina BDNF nel siero, che sembrano aumentare insieme al trattamento antidepressivo.68 La più recente di queste analisi suggerisce che queste aberrazioni del BDNF sono più pronunciate nei pazienti più gravemente depressi, ma che gli antidepressivi sembrano aumentare i livelli di questa proteina anche in assenza di remissione clinica.71 proBDNF è stato studiato meno ampiamente rispetto alla forma matura di BDNF, ma i due sembrano differire funzionalmente (in termini di effetti sui recettori della tirosina chinasi B) e recenti l'evidenza suggerisce che mentre il BDNF maturo può essere ridotto nella depressione, il proBDNF può essere sovrapprodotto.70 Il fattore di crescita nervoso valutato perifericamente è stato anche segnalato come inferiore nella depressione rispetto ai controlli in una meta-analisi, ma potrebbe non essere alterato dal trattamento antidepressivo nonostante sia più attenuato nei pazienti con depressione più grave.72 Risultati simili sono stati riportati in una meta-analisi per le cellule glialifattore neurotrofico derivato dalla linea 73
Il fattore di crescita dell'endotelio vascolare (VEGF) ha un ruolo nel promuovere l'angiogenesi e la neurogenesi insieme ad altri membri della famiglia VEGF (p. Es., VEGF-C, VEGF-D) e promette la depressione.75 Nonostante l'evidenza incoerente, due meta-analisi hanno aumenti recentemente indicati di VEGF nel sangue di pazienti depressi rispetto ai controlli (in 16 studi; P <0.001) .76,77 Tuttavia, un basso VEGF è stato identificato in TRD78 e livelli più alti hanno predetto la non risposta al trattamento antidepressivo.79 Non è chiaro perché i livelli di proteina VEGF sarebbero elevati, ma può essere in parte attribuibile all'attività proinfiammatoria e / o all'aumento della permeabilità della barriera ematoencefalica negli stati depressi che causa una ridotta espressione nel liquido cerebrospinale.80 La relazione tra VEGF e risposta al trattamento non è chiara. ; uno studio recente non ha rilevato alcuna relazione tra il VEGF sierico o il BDNF con la gravità della risposta o della depressione, nonostante le diminuzioni insieme al trattamento antidepressivo.81 Il fattore di crescita insulino-simile-1 è un fattore aggiuntivo con funzioni neurogeniche che possono essere aumentate nella depressione, riflettendo uno squilibrio nel processi neurotrofici.82,83 Il fattore di crescita dei fibroblasti di base (o FGF-2) è un membro della famiglia dei fattori di crescita dei fibroblasti e appare più elevato nei gruppi depressi rispetto ai gruppi di controllo.84 Tuttavia, i rapporti non sono coerenti; uno ha scoperto che questa proteina era più bassa nel MDD rispetto ai controlli sani, ma ridotta ulteriormente insieme al trattamento antidepressivo
Ulteriori fattori di crescita che non sono stati sufficientemente esplorati in depressione includono la tirosina chinasi 2 e la solubile tirosina chinasi-1 (anche denominata sVEGFR-1) che agiscono in sinergia con VEGF e i recettori tirosin-chinasi (che legano il BDNF) possono essere attenuati in depressione.86 Il fattore di crescita placentare è anche parte della famiglia VEGF, ma non è stato studiato in campioni sistematicamente depressi a nostra conoscenza.
Risultati metabolici dei biomarker nella depressione
I principali biomarker associati alla malattia metabolica comprendono leptina, adiponectina, grelina, trigliceridi, lipoproteine ad alta densità (HDL), glucosio, insulina e albumina.87 Le associazioni tra molti di questi e la depressione sono state esaminate: leptin88 e ghrelin89 appaiono inferiori nella depressione rispetto ai controlli nella periferia e possono aumentare insieme al trattamento antidepressivo o alla remissione. La resistenza all'insulina può essere aumentata in depressione, anche se con piccole quantità.90 I profili lipidici, compreso il colesterolo HDL, appaiono alterati in molti pazienti con depressione, compresi quelli senza comorbilità, sebbene questa relazione sia complessa e richieda ulteriori chiarimenti.91 Inoltre, iperglicemia92 e ipoalbuminemia93 in depressione sono stati riportati nelle revisioni.
Le indagini sugli stati metabolici generali stanno diventando più frequenti utilizzando pannelli metabolomici di piccole molecole con la speranza di trovare una solida firma biochimica per i disturbi psichiatrici. In un recente studio che utilizzava modelli di intelligenza artificiale, una serie di metaboliti che illustravano un aumento della segnalazione del glucosio lipidico era altamente predittiva di una diagnosi di MDD, 94 a sostegno di studi precedenti.95
Risultati del neurotrasmettitore nella depressione
Sebbene l'attenzione prestata alle monoamine nella depressione abbia prodotto trattamenti di relativamente successo, non sono stati identificati marcatori di neurotrasmettitori robusti per ottimizzare il trattamento basato sulla selettività dei bersagli monoaminici degli antidepressivi. Lavori recenti indicano che il recettore 5A della serotonina (1-idrossitriptamina) è potenzialmente importante sia per la diagnosi che per la prognosi della depressione, in attesa di nuove tecniche genetiche e di imaging.96 Esistono nuovi potenziali trattamenti mirati alla 5-idrossitriptamina; per esempio, utilizzando una somministrazione a rilascio lento di 5-idrossitriptofano.97 L'aumentata trasmissione della dopamina interagisce con altri neurotrasmettitori per migliorare i risultati cognitivi come il processo decisionale e la motivazione.98 Allo stesso modo, i neurotrasmettitori glutammato, noradrenalina, istamina e serotonina possono interagire e attivarsi come parte di una risposta allo stress correlata alla depressione; questo potrebbe diminuire la produzione di 5-idrossitriptamina attraverso le `` inondazioni ''. Una recente revisione espone questa teoria e suggerisce che nella TRD, questa potrebbe essere invertita (e ripristinata la 5-HT) attraverso un trattamento multimodale mirato a più neurotrasmettitori.99 È interessante notare che gli aumenti della serotonina non si verificano sempre insieme ai benefici terapeutici degli antidepressivi.100 Nonostante ciò , è stato spesso riscontrato che i metaboliti dei neurotrasmettitori come il 3-metossi-4-idrossifenilglicole, della noradrenalina o l'acido omovanillico della dopamina aumentano parallelamente alla riduzione della depressione con il trattamento antidepressivo101,102 o che bassi livelli di questi metaboliti predicono una migliore risposta a Trattamento SSRI 102,103
Risultati neuroendocrini nella depressione
Il cortisolo è il più comune biomarker dell'asse HPA studiato in depressione. Numerose recensioni si sono concentrate sulle varie valutazioni dell'attività di HPA; nel complesso, questi indicano che la depressione è associata all'ipercortisolemia e che la risposta al risveglio del cortisolo è spesso attenuata. 104,105 Ciò è supportato da una recente revisione dei livelli cronici di cortisolo misurati nei capelli, supportando l'iperattività del cortisolo in depressione ma l'ipoattività in altre malattie come come disturbo di panico.106 Inoltre, in particolare, livelli elevati di cortisolo possono predire una risposta più scarsa al trattamento psicologico107 e antidepressivo di 108. Storicamente, il marker neuroendocrino più promettente della risposta al trattamento prospettico è stato il test di soppressione del desametasone, in cui la non soppressione del cortisolo in seguito alla somministrazione di desametasone è associata a una minore probabilità di successiva remissione. Tuttavia, questo fenomeno non è stato considerato sufficientemente robusto per l'applicazione clinica. I marcatori correlati, l'ormone rilasciante corticotropina e l'ormone adrenocorticotropina, nonché la vasopressina, sono stati trovati in modo incoerente come sovrapprodotti nella depressione e il deidroepiandrosterone è risultato attenuato; il rapporto tra cortisolo e deidroepiandrosterone può essere elevato come marker relativamente stabile nella TRD, persistendo dopo la remissione.109 Le disfunzioni dell'ormone neuroendocrino sono state a lungo associate alla depressione e l'ipotiroidismo può anche svolgere un ruolo causale nell'umore depresso.110 Inoltre, le risposte della tiroide possono normalizzare con successo il trattamento per la depressione.111
All'interno di quanto sopra, è importante anche considerare le vie di segnalazione attraverso i sistemi, come la glicogeno sintasi chinasi-3, la proteina chinasi attivata da mitogeno e l'adenosina ciclica 3?, 5? -Monofosfato, coinvolti nella plasticità sinaptica112 e modificati da antidepressivi.113 Ulteriori informazioni potenziali candidati biomarcatori che abbracciano i sistemi biologici in particolare sono misurati utilizzando neuroimaging o genetica. In risposta alla mancanza di differenze genomiche robuste e significative tra popolazioni depresse e non depresse, 114 nuovi approcci genetici come i punteggi poligenici115 o la lunghezza dei telomeri116,117 potrebbero rivelarsi più utili. Ulteriori biomarcatori che stanno guadagnando popolarità stanno esaminando i cicli circadiani oi biomarcatori cronobiologici utilizzando diverse fonti. L'actigrafia può fornire una valutazione oggettiva dell'attività del sonno e della veglia e del riposo attraverso un accelerometro, ei dispositivi actigrafici possono misurare sempre più fattori aggiuntivi come l'esposizione alla luce. Questo può essere più utile per il rilevamento rispetto ai rapporti soggettivi di pazienti comunemente usati e potrebbe fornire nuovi predittori della risposta al trattamento.118 La questione di quali biomarcatori siano i più promettenti per l'uso traslazionale è impegnativa, che viene ampliata di seguito.
Sfide attuali
Per ciascuno di questi cinque sistemi neurobiologici esaminati, le prove seguono una narrativa simile: ci sono molti biomarcatori che esistono associati per alcuni aspetti alla depressione. Questi indicatori sono spesso correlati in modo complesso e difficile da modello. L'evidenza è incoerente ed è probabile che alcuni siano epifenomeni di altri fattori e alcuni siano importanti solo in un sottogruppo di pazienti. È probabile che i biomarcatori siano utili attraverso una varietà di percorsi (ad esempio quelli che predicono la successiva risposta al trattamento, quelli che indicano trattamenti specifici come più efficaci o quelli che si modificano con interventi indipendentemente dai miglioramenti clinici). Sono necessari nuovi metodi per massimizzare la coerenza e l'applicabilità clinica delle valutazioni biologiche nelle popolazioni psichiatriche.
Variabilità dei biomarcatori
La variazione dei biomarcatori nel tempo e in tutte le situazioni riguarda più alcuni tipi (ad esempio la proteomica) rispetto ad altri (la genomica). Le norme standardizzate per molti non esistono o non sono state ampiamente accettate. In effetti, l'influenza dei fattori ambientali sui marcatori dipende spesso dalla composizione genetica e da altre differenze fisiologiche tra le persone che non possono essere tutte spiegate. Ciò rende la valutazione dell'attività dei biomarcatori e l'identificazione di anomalie biologiche, difficili da interpretare. A causa del numero di potenziali biomarcatori, molti non sono stati misurati ampiamente o in un pannello completo insieme ad altri marcatori rilevanti.
Molti fattori sono stati segnalati per alterare i livelli di proteine nei sistemi biologici in pazienti con disturbi affettivi. Insieme a fattori correlati alla ricerca come la durata e le condizioni di conservazione (che possono causare la degradazione di alcuni composti), questi includono il tempo del giorno misurato, l'etnia, l'esercizio fisico, la dieta 119 (ad esempio, l'attività del microbioma, specialmente a condizione che la maggior parte degli studi sui biomarcatori del sangue facciano non richiede un campione di digiuno), fumo 120 e uso di sostanze, 121 così come fattori di salute (come malattie infiammatorie comorbili, cardiovascolari o di altro tipo). Ad esempio, sebbene l'aumento dell'infiammazione sia osservato negli individui depressi ma altrimenti sani rispetto ai gruppi non depressi, gli individui depressi che hanno anche una condizione di comorbilità immunitaria spesso hanno livelli ancora più elevati di citochine rispetto a quelli senza depressione o malattia.122 Alcuni fattori importanti con Il probabile coinvolgimento nella relazione tra biomarcatori, depressione e risposta al trattamento sono descritti di seguito.
Stress. Entrambe le risposte endocrine e immunitarie hanno ruoli ben noti nel rispondere allo stress (fisiologico o psicologico) e lo stress transitorio al momento della raccolta dei campioni biologici è raramente misurato negli studi di ricerca, nonostante la variabilità di questo fattore tra individui che può essere accentuata dagli attuali sintomi depressivi. Sia gli stress psicologici acuti che quelli cronici agiscono come una sfida immunitaria, accentuando le risposte infiammatorie a breve e lungo termine.123,124 Questa scoperta si estende all'esperienza dello stress precoce, che è stato associato a aumenti infiammatori adulti che sono indipendenti dallo stress sperimentato come un adulto.125,126 Durante l'esperienza traumatica infantile, è stata riportata anche un'infiammazione intensificata solo in quei bambini che erano attualmente depressi.127 Al contrario, le persone con depressione e una storia di trauma infantile possono avere risposte cortisoliche allo stress, rispetto a quelle con depressione e nessun trauma nella prima infanzia.128 Le alterazioni dell'asse HPA indotte dallo stress appaiono correlate con la funzione cognitiva, 129 così come il sottotipo di depressione o variazione nei geni correlati a HPA.130 Lo stress ha anche effetti di compromissione a breve e lungo termine sulla neurogenesi131 e altri neuroni meccanismi.132 Non è chiaro esattamente come il trauma infantile influisca sui marcatori biologici in depressione adulti, ma è possibile che lo stress precoce predisponga alcuni individui a sopportare reazioni di stress nell'età adulta amplificate psicologicamente e / o biologicamente.
Funzionamento cognitivo. Le disfunzioni neurocognitive si verificano frequentemente nelle persone con disturbi affettivi, anche in MDD.133 non-diagnosticato. I deficit cognitivi appaiono cumulativi accanto alla resistenza al trattamento. 134 Neurobiologicamente, l'asse HPXXUMX e i sistemi neurotrofici 129 giocheranno probabilmente un ruolo chiave in questa relazione. I neurotrasmettitori noradrenalina e dopamina sono probabilmente importanti per i processi cognitivi come l'apprendimento e la memoria.135 Elevate risposte infiammatorie sono state collegate al declino cognitivo e probabilmente influenzano il funzionamento cognitivo negli episodi depressivi, 136 e in remissione, attraverso una varietà di meccanismi.137 Infatti, Krogh e al138 hanno proposto che la CRP sia più strettamente correlata alle prestazioni cognitive rispetto ai sintomi principali della depressione.
Età, sesso e indice di massa corporea. L'assenza o la presenza e la direzione delle differenze biologiche tra uomini e donne sono state particolarmente variabili nelle prove fino ad oggi. La variazione dell'ormone neuroendocrino tra uomini e donne interagisce con la suscettibilità alla depressione.140 Una revisione degli studi sull'infiammazione ha riportato che il controllo per età e sesso non ha influenzato le differenze di controllo del paziente nelle citochine infiammatorie (sebbene l'associazione tra IL-6 e depressione si riducesse con l'aumentare dell'età, che è coerente con le teorie secondo le quali l'infiammazione aumenta generalmente con l'età) .41,141 Le differenze di VEGF tra pazienti e controlli sono maggiori negli studi che valutano i campioni più giovani, mentre il sesso, l'IMC ei fattori clinici non hanno influenzato questi confronti a livello meta-analitico. la mancanza di aggiustamento per l'IMC in precedenti esami di infiammazione e depressione sembra confondere le differenze altamente significative riportate tra questi gruppi.77 Il tessuto adiposo allargato è stato definitivamente dimostrato di stimolare la produzione di citochine e di essere strettamente legato ai marcatori metabolici.41 Perché i farmaci psicotropi può essere associato con un aumento di peso e un IMC più elevato, e questi sono stati associati alla resistenza al trattamento in depressione, questa è un'area importante da esaminare.
Farmaci. Molti studi sui biomarcatori nella depressione (sia trasversali che longitudinali) hanno raccolto campioni di riferimento in partecipanti non medicati per ridurre l'eterogeneità. Tuttavia, molte di queste valutazioni vengono prese dopo un periodo di wash-out dal farmaco, che lascia il fattore di confusione potenzialmente significativo dei cambiamenti residui nella fisiologia, esacerbato dall'ampia gamma di trattamenti disponibili che potrebbero aver avuto effetti diversi sull'infiammazione. Alcuni studi hanno escluso l'uso di farmaci psicotropi, ma non di altri farmaci: in particolare, la pillola contraccettiva orale è frequentemente consentita nei partecipanti alla ricerca e non controllata nelle analisi, che è stato recentemente indicato per aumentare i livelli di ormoni e citochine.143,144 Diversi studi indicano che l'antidepressivo i farmaci hanno effetti sulla risposta infiammatoria, 34,43,49,145 147 asse HPA, 108 neurotrasmettitore, 148 e attività neurotrofica149. Tuttavia, i numerosi potenziali trattamenti per la depressione hanno proprietà farmacologiche distinte e complesse, suggerendo che potrebbero esserci effetti biologici discreti di diverse opzioni di trattamento, supportati dai dati attuali. È stato teorizzato che, oltre agli effetti delle monoamine, è probabile che specifici farmaci mirati alla serotonina (cioè gli SSRI) prendano di mira i cambiamenti Th2 nell'infiammazione e gli antidepressivi noradrenergici (p.es., gli SNRI) influenzino un cambiamento Th1 Non è ancora possibile determinare gli effetti di farmaci singoli o combinati sui biomarcatori. Questi sono probabilmente mediati da altri fattori tra cui la durata del trattamento (pochi studi valutano l'uso di farmaci a lungo termine), l'eterogeneità del campione e la non stratificazione dei partecipanti in base alla risposta al trattamento.
Eterogeneità
Metodologico. Come accennato in precedenza, le differenze (tra e all'interno degli studi) in termini di quali terapie (e combinazioni) i partecipanti stanno prendendo e hanno preso in precedenza sono destinate a introdurre eterogeneità nei risultati della ricerca, in particolare nella ricerca sui biomarcatori. Oltre a questo, molte altre caratteristiche di progettazione e campionatura variano tra gli studi, aumentando così la difficoltà di interpretare e attribuire risultati. Questi includono i parametri di misurazione dei biomarker (ad es. Kit di dosaggio) e metodi di raccolta, conservazione, elaborazione e analisi dei marker in depressione. Hiles et al141 hanno esaminato alcune fonti di incoerenza nella letteratura sull'infiammazione e hanno trovato che l'accuratezza della diagnosi di depressione, BMI e comorbidità erano i più importanti da considerare nella valutazione dell'infiammazione periferica tra gruppi depressi e non depressi.
Clinico. L'ampia eterogeneità delle popolazioni depresse è ben documentata con151 ed è un contributo fondamentale ai risultati contrastanti all'interno della letteratura di ricerca. È probabile che, anche all'interno delle diagnosi, profili biologici anormali siano limitati a sottoinsiemi di individui che potrebbero non essere stabili nel tempo. Sottogruppi coesivi di persone che soffrono di depressione possono essere identificabili attraverso una combinazione di fattori psicologici e biologici. Di seguito, delineiamo il potenziale per esplorare i sottogruppi nel soddisfare le sfide poste dalla variabilità dei biomarcatori e dall'eterogeneità.
Sottotipi all'interno della depressione
Finora, nessun sottogruppo omogeneo all'interno di episodi o disturbi di depressione è stato in grado di distinguere in modo affidabile tra i pazienti in base alla presentazione dei sintomi o alla risposta al trattamento.152 L'esistenza di un sottogruppo in cui le aberrazioni biologiche sono più pronunciate aiuterebbe a spiegare l'eterogeneità tra studi precedenti e potrebbe catalizzare il percorso verso il trattamento stratificato. Kunugi et al153 hanno proposto una serie di quattro potenziali sottotipi basati sul ruolo di diversi sistemi neurobiologici che mostrano sottotipi clinicamente rilevanti nella depressione: quelli con ipercortisolismo che si presenta con depressione malinconica, o ipocortisolismo che riflette un sottotipo atipico, un sottogruppo di pazienti correlato alla dopamina che può presente in modo prominente con l'anedonia (e potrebbe rispondere bene, ad esempio, all'aripiprazolo) e un sottotipo infiammatorio caratterizzato da un'infiammazione elevata. Molti articoli incentrati sull'infiammazione hanno specificato il caso dell'esistenza di un `` sottotipo infiammatorio '' all'interno della depressione.55,56,154,155 I correlati clinici di un'infiammazione elevata sono ancora indeterminati e sono stati fatti pochi tentativi diretti per scoprire quali partecipanti possono comprendere questa coorte. È stato proposto che le persone con depressione atipica possano avere livelli di infiammazione più elevati rispetto al sottotipo malinconico, 156, il che forse non è in linea con i risultati relativi all'asse HPA nei sottotipi di depressione malinconici e atipici. Anche TRD37 o depressione con sintomi somatici prominenti157 è stato ipotizzato come potenziale sottotipo infiammatorio, ma neurovegetativo (sonno, appetito, perdita di libido), umore (incluso umore basso, tendenza al suicidio e irritabilità) e sintomi cognitivi (inclusi pregiudizi affettivi e senso di colpa) 158 tutti sembrano correlati a profili biologici. Ulteriori potenziali candidati per un sottotipo infiammatorio riguardano l'esperienza di sintomi simili al comportamento della malattia159,160 o una sindrome metabolica.158
La propensione verso (ipo) mania può distinguere biologicamente tra i pazienti che soffrono di depressione. Le prove ora suggeriscono che le malattie bipolari sono un gruppo sfaccettato di disturbi dell'umore, con il disturbo bipolare subsindromale riscontrato in modo più prevalente di quanto precedentemente riconosciuto. 161 L'individuazione imprecisa e / o ritardata del disturbo bipolare è stata recentemente evidenziata come un problema importante nella psichiatria clinica, con tempo medio per correggere la diagnosi frequentemente superando un decennio162 e questo ritardo causa una maggiore gravità e costo della malattia generale. 163 Con la maggior parte dei pazienti con disturbo bipolare che si presentano inizialmente con uno o più episodi depressivi e depressione unipolare è la diagnosi errata più frequente, l'identificazione di fattori che potrebbero differenziare tra depressione unipolare e bipolare ha implicazioni sostanziali.164 I disturbi dello spettro bipolare probabilmente non sono stati rilevati in alcune precedenti indagini sui biomarker MDD, e l'infusione di evidenze ha indicato la differenziazione dell'attività dell'asse HPA109 o l'infiammazione165,166 tra bipolare e unipo depressione Tuttavia, questi confronti sono scarsi, possiedono piccole dimensioni del campione, identificati effetti di trend non significativi o popolazioni reclutate che non erano ben caratterizzate dalla diagnosi. Queste indagini inoltre non esaminano il ruolo della risposta al trattamento in queste relazioni.
Entrambi i disturbi bipolari167 e la resistenza al trattamento168 non sono costrutti dicotomici e si trovano su continui, il che aumenta la sfida dell'identificazione del sottotipo. Oltre alla sottotipizzazione, vale la pena notare che molte anomalie biologiche osservate nella depressione sono analogamente riscontrate in pazienti con altre diagnosi. Pertanto, gli esami diagnostici transdermici sono anche potenzialmente importanti.
Sfide di misurazione dei biomarcatori
Selezione di biomarcatori. Il gran numero di biomarcatori potenzialmente utili rappresenta una sfida per la psicobiologia nel determinare quali marcatori sono implicati in che modo e per chi. Per aumentare la sfida, relativamente pochi di questi biomarcatori sono stati oggetto di indagini sufficienti sulla depressione e, per la maggior parte, i loro ruoli precisi nelle popolazioni sane e cliniche non sono ben compresi. Nonostante ciò, sono stati fatti numerosi tentativi per proporre promettenti pannelli di biomarcatori. Oltre a 16 serie di marcatori di Brand et al. Con un forte potenziale, 27 Lopresti et al. Delineano un ulteriore ampio set di marcatori di stress ossidativo con potenzialità per migliorare la risposta al trattamento.28 Papakostas et al. Hanno definito a priori una serie di nove marcatori sierici che si estendono sistemi biologici (BDNF, cortisolo, recettore solubile del TNF? di tipo II, alfa1 antitripsina, apolipoproteina CIII, fattore di crescita epidermico, mieloperossidasi, prolattina e resistina) in campioni di validazione e replicazione con MDD. Una volta combinati, una misura composita di questi livelli è stata in grado di distinguere tra MDD e gruppi di controllo con un'accuratezza dell'80% -90 %.169 Proponiamo che anche questi non coprano tutti i potenziali candidati in questo campo; vedere la Tabella 2 per una delineazione non esaustiva dei biomarcatori con potenziale depressione, contenente sia quelli con una base di prove sia promettenti nuovi marcatori.
Tecnologia. A causa dei progressi tecnologici, è ora possibile (anzi, conveniente) misurare una vasta gamma di biomarcatori contemporaneamente a un costo inferiore e con una sensibilità maggiore rispetto a prima. Al momento, questa capacità di misurare numerosi composti è in anticipo sulla nostra capacità di analizzare e interpretare efficacemente i dati, 170 qualcosa che continuerà con l'aumento degli array di biomarcatori e di nuovi marcatori come con la metabolomica. Ciò è in gran parte dovuto alla mancanza di comprensione circa i ruoli precisi e le interrelazioni tra i marcatori e una comprensione insufficiente di come i marcatori correlati si associano a diversi livelli biologici (ad esempio, genetica, trascrizione, proteine) all'interno e tra individui. I grandi dati che utilizzano nuovi approcci e standard analitici aiuteranno a risolvere questo problema e verranno proposte nuove metodologie; un esempio è lo sviluppo di un approccio statistico basato su un'analisi basata sul flusso per scoprire nuovi potenziali marcatori metabolici basati sulle loro reazioni tra reti e integrare l'espressione genica con i dati del metabolita.171 Le tecniche di apprendimento automatico sono già applicate e aiuteranno i modelli che utilizzano biomarker dati per prevedere i risultati del trattamento in studi con big data.172
Biomarcatori aggreganti. L'esame simultaneo di una serie di biomarcatori è un'alternativa all'ispezione di marcatori isolati che potrebbero fornire un punto di vista più accurato nella complessa rete di sistemi o reti biologici.26 Inoltre, per aiutare a districare le prove contrastanti in questa letteratura fino ad oggi (in particolare, dove le reti di biomarcatori e le interazioni sono ben comprese), i dati dei biomarcatori possono quindi essere aggregati o indicizzati. Una sfida consiste nell'identificare il metodo ottimale per condurre questa operazione e potrebbe richiedere miglioramenti nella tecnologia e / o nuove tecniche analitiche (vedere la sezione "Big data"). Storicamente, i rapporti tra due biomarcatori distinti hanno prodotto risultati interessanti.109,173 Sono stati fatti pochi tentativi per aggregare i dati dei biomarcatori su scala più ampia, come quelli che utilizzano l'analisi dei componenti principali delle reti di citochine proinfiammatorie.174 In una meta-analisi, le citochine proinfiammatorie sono state convertito in un punteggio di dimensione a effetto singolo per ogni studio, e nel complesso ha mostrato un'infiammazione significativamente più alta prima del trattamento antidepressivo, prevedendo la successiva non risposta negli studi ambulatoriali. I pannelli di biomarcatori compositi sono sia una sfida che un'opportunità per la ricerca futura per identificare risultati significativi e affidabili che possono essere applicati per migliorare i risultati del trattamento.43 Uno studio di Papakostas et al ha adottato un approccio alternativo, selezionando un pannello di biomarcatori sierici eterogenei (di infiammatori, Asse HPA e sistemi metabolici) che in uno studio precedente avevano indicato differivano tra individui depressi e di controllo e li avevano composti in un punteggio di rischio che differiva in due campioni indipendenti e in un gruppo di controllo con sensibilità e specificità> 80 %.169
Grandi dati L'uso di big data è probabilmente necessario per affrontare le attuali sfide delineate intorno all'eterogeneità, alla variabilità dei biomarcatori, identificare i marcatori ottimali e portare il campo verso la ricerca traslazionale applicata sulla depressione. Tuttavia, come indicato sopra, ciò comporta sfide tecnologiche e scientifiche.175 Le scienze della salute hanno iniziato a utilizzare l'analisi dei big data solo di recente, circa un decennio più tardi rispetto al settore aziendale. Tuttavia, studi come iSPOT-D152 e consorzi come lo Psychiatric Genetics Consortium176 stanno progredendo con la nostra comprensione dei meccanismi biologici in psichiatria. Gli algoritmi di apprendimento automatico hanno, in pochissimi studi, iniziato ad essere applicati ai biomarcatori per la depressione: una recente indagine ha raccolto dati da> 5,000 partecipanti di 250 biomarcatori; dopo molteplici imputazioni di dati, è stata condotta una regressione potenziata dall'apprendimento automatico, indicando 21 potenziali biomarcatori. A seguito di ulteriori analisi di regressione, sono stati selezionati tre biomarcatori che si associavano più fortemente ai sintomi depressivi (dimensioni dei globuli rossi altamente variabili, livelli sierici di glucosio e bilirubina). Gli autori concludono che i big data possono essere utilizzati efficacemente per generare ipotesi.177 Sono attualmente in corso progetti di fenotipizzazione di biomarcatori più grandi che aiuteranno a far avanzare il nostro viaggio nel futuro della neurobiologia della depressione.
Prospettive future
Identificazione del pannello dei biomarker
I risultati in letteratura fino ad oggi richiedono la replica in studi su larga scala. Ciò è particolarmente vero per i nuovi biomarcatori, come la chemochina timo e la chemochina regolata dall'attivazione e il fattore di crescita tirosin chinasi 2 che, a nostra conoscenza, non sono stati studiati in campioni di controllo clinicamente depressi e sani. Gli studi sui big data devono analizzare pannelli di biomarcatori completi e utilizzare tecniche di analisi sofisticate per accertare appieno le relazioni tra i marcatori e quei fattori che li modificano nelle popolazioni cliniche e non cliniche. Inoltre, repliche su larga scala dell'analisi delle componenti principali potrebbero stabilire gruppi altamente correlati di biomarcatori e potrebbero anche informare l'uso di `` composti '' nella psichiatria biologica, il che potrebbe migliorare l'omogeneità dei risultati futuri.
Scoperta di sottotipi omogenei
Per quanto riguarda la selezione dei biomarker, potrebbero essere necessari più pannelli per diversi potenziali percorsi che la ricerca potrebbe implicare. Nel loro insieme, le prove attuali indicano che i profili di biomarcatori sono sicuramente, ma astrusi in una sottopopolazione di individui che attualmente soffrono di depressione. Questo può essere stabilito all'interno o attraverso categorie diagnostiche, il che renderebbe conto di alcune incoerenze delle scoperte che possono essere osservate in questa letteratura. La quantificazione di un sottogruppo biologico (o sottogruppi) può essere più efficacemente facilitata da una grande analisi a grappolo di pannelli di rete di biomarker nella depressione. Ciò illustrerebbe la variabilità all'interno della popolazione; analisi della classe latente potrebbero presentare caratteristiche cliniche distinte basate, ad esempio, sull'infiammazione.
Effetti specifici del trattamento su infiammazione e risposta
Tutti i trattamenti comunemente prescritti per la depressione dovrebbero essere valutati in modo completo per i loro specifici effetti biologici, tenendo conto anche dell'efficacia delle prove di trattamento. Ciò potrebbe consentire a costrutti relativi ai biomarcatori e alle presentazioni di sintomi di predire i risultati di una varietà di trattamenti antidepressivi in modo più personalizzato e potrebbe essere possibile nel contesto della depressione unipolare e bipolare. Questo è probabilmente utile per nuovi trattamenti potenziali e trattamenti attualmente indicati.
Determinazione prospettica della risposta al trattamento
L'uso delle tecniche di cui sopra può portare a una migliore capacità di prevedere la resistenza del trattamento in modo prospettico. Misure più autentiche e persistenti (ad esempio, a lungo termine) della risposta al trattamento possono contribuire a questo. La valutazione di altre misure valide per il benessere del paziente (come la qualità della vita e il funzionamento quotidiano) potrebbe fornire una valutazione più olistica dell'esito del trattamento che potrebbe associarsi più strettamente ai biomarcatori. Mentre l'attività biologica da sola potrebbe non essere in grado di distinguere i risponditori del trattamento da non responders, la misurazione concomitante di biomarcatori con variabili psicosociali o demografiche potrebbe essere integrata con informazioni sui biomarcatori nello sviluppo di un modello predittivo di risposta al trattamento insufficiente. Se un modello affidabile viene sviluppato per prevedere la risposta (sia per la popolazione depressa che per una sottopopolazione) ed è validato in modo retrospettivo, un progetto di traduzione può stabilire la sua applicabilità in un ampio studio controllato.
Verso trattamenti stratificati
Al momento, i pazienti con depressione non sono sistematicamente diretti a ricevere un programma di intervento ottimizzato. Se convalidato, un disegno di prova stratificato potrebbe essere impiegato per testare un modello per prevedere la non risposta e / o per determinare dove un paziente deve essere valutato in un modello di cura a gradini. Questo potrebbe essere utile sia in contesti di trattamento standardizzati che naturalistici, attraverso diversi tipi di intervento. Infine, potrebbe essere sviluppato un modello clinicamente valido per fornire agli individui il trattamento più appropriato, per riconoscere coloro che hanno probabilità di sviluppare depressione refrattaria e fornire cure e monitoraggio migliori a questi pazienti. Ai pazienti identificati come a rischio di resistenza al trattamento può essere prescritta una terapia psicologica e farmacologica concomitante o una farmacoterapia combinata. A titolo di esempio speculativo, i partecipanti senza aumenti delle citochine proinfiammatorie potrebbero essere indicati per ricevere una terapia psicologica piuttosto che farmacologica, mentre un sottogruppo di pazienti con infiammazione particolarmente alta potrebbe ricevere un agente antinfiammatorio in aggiunta al trattamento standard. Analogamente alla stratificazione, in futuro potrebbero essere possibili strategie di selezione del trattamento personalizzate. Ad esempio, un particolare individuo depresso potrebbe avere un TNF marcatamente alto? livelli, ma nessun'altra anomalia biologica, e potrebbe beneficiare di un trattamento a breve termine con un TNF? antagonista.54 Il trattamento personalizzato può anche comportare il monitoraggio dell'espressione del biomarcatore durante il trattamento per informare su possibili modifiche dell'intervento, sulla durata della terapia di continuazione richiesta o per rilevare i marker precoci di ricaduta.
Nuovi obiettivi di trattamento
Esiste un numero enorme di potenziali trattamenti che potrebbero essere efficaci per la depressione, che non sono stati adeguatamente esaminati, inclusi interventi nuovi o riproposti da altre discipline mediche. Alcuni degli obiettivi più popolari sono stati i farmaci antinfiammatori come il celecoxib (e altri inibitori della cicloossigenasi-2), il TNF? antagonisti etanercept e infliximab, minociclina o aspirina. Questi sembrano promettenti.178 I composti antiglucocorticoidi, tra cui ketoconazolo179 e metirapone, 180 sono stati studiati per la depressione, ma entrambi hanno svantaggi con il loro profilo di effetti collaterali e il potenziale clinico del metirapone è incerto. Anche il mifepristone181 ei corticosteroidi fludrocortisone e spironolattone 182 e il desametasone e l'idrocortisone183 possono essere efficaci nel trattamento della depressione a breve termine. Mirare agli antagonisti del recettore del glutammato N-metil-d-aspartato, inclusa la ketamina, potrebbe rappresentare trattamenti efficaci nella depressione.184 Gli acidi grassi polinsaturi omega-3 influenzano l'attività infiammatoria e metabolica e sembrano dimostrare una certa efficacia per la depressione.185 È possibile che le statine possano hanno effetti antidepressivi186 attraverso percorsi neurobiologici rilevanti.187
In questo modo, gli effetti biochimici degli antidepressivi (vedere la sezione `` Farmaci '') sono stati utilizzati per benefici clinici in altre discipline: in particolare malattie sintomatiche gastroenterologiche, neurologiche e non specifiche.188 Gli effetti antinfiammatori degli antidepressivi possono rappresentare parte del meccanismo di questi vantaggi. È stato anche suggerito che il litio riduca l'infiammazione, in modo critico attraverso i percorsi della glicogeno sintasi chinasi-3 Un focus su questi effetti potrebbe rivelarsi informativo per la firma di un biomarcatore della depressione e, a loro volta, i biomarcatori potrebbero rappresentare marcatori surrogati per lo sviluppo di nuovi farmaci.
Insight di Dr. Alex Jimenez
La depressione è una malattia mentale caratterizzata da gravi sintomi che influenzano l'umore, compresa la perdita di interesse nelle attività. Recenti studi di ricerca, tuttavia, hanno scoperto che potrebbe essere possibile diagnosticare la depressione utilizzando più di semplici sintomi comportamentali di un paziente. Secondo i ricercatori, l'identificazione di biomarcatori facilmente ottenibili che potrebbero diagnosticare in modo più accurato la depressione è fondamentale per migliorare la salute e il benessere generale di un paziente. A titolo di esempio, i risultati clinici suggeriscono che i soggetti con disturbo depressivo maggiore, o MDD, hanno livelli più bassi della molecola di acetil-L-carnitina, o LAC, nel sangue rispetto ai controlli sani. In definitiva, stabilire biomarcatori per la depressione potrebbe potenzialmente aiutare a determinare meglio chi è a rischio di sviluppare il disturbo e aiutare gli operatori sanitari a determinare la migliore opzione di trattamento per un paziente con depressione.
Conclusione
La letteratura indica che circa i due terzi dei pazienti con depressione non raggiungono la remissione a un trattamento iniziale e che la probabilità di mancata risposta aumenta con il numero di trattamenti sperimentati. Fornire terapie inefficaci ha conseguenze sostanziali per i costi individuali e sociali, tra cui sofferenza persistente e scarso benessere, rischio di suicidio, perdita di produttività e sprechi di risorse sanitarie. La vasta letteratura in depressione indica un numero enorme di biomarcatori con il potenziale per migliorare il trattamento per le persone con depressione. Oltre ai neurotrasmettitori e ai marcatori neuroendocrini che sono stati oggetto di studio diffuso per molti decenni, recenti intuizioni evidenziano la risposta infiammatoria (e il sistema immunitario più in generale), i fattori metabolici e di crescita come importanti coinvolti nella depressione. Tuttavia, prove contrastanti e eccessive dimostrano che è necessario affrontare una serie di sfide prima di poter applicare la ricerca sui biomarcatori al fine di migliorare la gestione e la cura delle persone depresse. A causa della pura complessità dei sistemi biologici, gli esami simultanei di una gamma completa di marcatori in campioni di grandi dimensioni sono di notevole utilità nello scoprire interazioni tra stati biologici e psicologici tra individui. È probabile che l'ottimizzazione della misurazione dei parametri neurobiologici e delle misure cliniche della depressione faciliti una maggiore comprensione. Questa recensione evidenzia anche l'importanza di esaminare i fattori potenzialmente modificanti (come la malattia, l'età, la cognizione e le terapie) nello spianare una comprensione coerente della biologia della depressione e dei meccanismi di resistenza del trattamento. È probabile che alcuni marcatori mostrino la massima promessa di prevedere la risposta al trattamento o la resistenza a trattamenti specifici in un sottogruppo di pazienti, e la misurazione concomitante di dati biologici e psicologici può migliorare la capacità di identificare prospetticamente i soggetti a rischio per esiti negativi del trattamento. L'istituzione di un panel di biomarcatori ha implicazioni per aumentare l'accuratezza diagnostica e la prognosi, nonché per individualizzare i trattamenti nella fase più precoce possibile della malattia depressiva e sviluppare nuovi efficaci bersagli terapeutici. Queste implicazioni possono essere limitate a sottogruppi di pazienti depressi. I percorsi verso queste possibilità integrano le recenti strategie di ricerca per collegare le sindromi cliniche più da vicino ai substrati neurobiologici sottostanti.6 Oltre a ridurre l'eterogeneità, ciò può facilitare uno spostamento verso la parità di stima tra salute fisica e mentale. È chiaro che, sebbene sia necessario molto lavoro, l'istituzione della relazione tra biomarker rilevanti e disturbi depressivi ha implicazioni sostanziali per ridurre il peso della depressione a livello individuale e sociale.
Ringraziamenti
Questo rapporto rappresenta una ricerca indipendente finanziata dal National Institute for Health Research (NIHR) Biomedical Research Center a South London e Maudsley NHS Foundation Trust e King's College London. Le opinioni espresse sono quelle degli autori e non necessariamente quelle del NHS, del NIHR o del Dipartimento della Salute.
Le note
Rivelazione. AHY ha ricevuto negli ultimi 3 anni onorari per aver parlato con Astra Zeneca (AZ), Lundbeck, Eli Lilly, Sunovion; onorario per la consulenza di Allergan, Livanova e Lundbeck, Sunovion, Janssen; e sostegno alla ricerca da parte di Janssen e delle agenzie di finanziamento del Regno Unito (NIHR, MRC, Wellcome Trust). Negli ultimi 3, AJC ha ricevuto onorari per aver parlato con Astra Zeneca (AZ), onorario per la consulenza di Allergan, Livanova e Lundbeck, e il sostegno alla ricerca da parte di Lundbeck e delle agenzie di finanziamento del Regno Unito (NIHR, MRC, Wellcome Trust).
Gli autori non segnalano altri conflitti di interesse in questo lavoro.
In conclusione,Mentre numerosi studi di ricerca hanno trovato centinaia di biomarcatori per la depressione, non molti hanno stabilito il loro ruolo nella malattia depressiva o come potrebbero essere utilizzate esattamente informazioni biologiche per migliorare la diagnosi, il trattamento e la prognosi. Tuttavia, l'articolo di cui sopra esamina la letteratura disponibile sui biomarcatori coinvolti durante altri processi e confronta i risultati clinici con quelli della depressione. Inoltre, nuove scoperte sui biomarcatori per la depressione possono aiutare a diagnosticare meglio la depressione al fine di seguire un trattamento migliore. Informazioni referenziate dal National Center for Biotechnology Information (NCBI) . L'ambito delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
1. Prince M, Patel V, Saxena S, et al. Nessuna salute senza salute mentaleLancet. 2007;370(9590): 859.[PubMed]
2. Kingdon D, Wykes T. Aumento dei fondi necessari per la ricerca sulla salute mentaleBMJ. 2013;346: F402.[PubMed]
3. Vivekanantham S, Strawbridge R, Rampuri R, Ragunathan T, Young AH. Parità di pubblicazione per la psichiatriaBr J Psychiatry. 2016;209(3): 257 261. [PubMed]
4. Fava M. Diagnosi e definizione di depressione resistente al trattamentoBiol Psychiatry. 2003;53(8): 649 659. [PubMed]
5. Insel T, Cuthbert B, Garvey M, et al. Criteri di dominio della ricerca (RDoC): verso un nuovo quadro di classificazione per la ricerca sui disturbi mentaliSono J Psychiatry. 2010;167(7): 748 751. [PubMed]
6. Kapur S, Phillips AG, Insel TR. Perché ci è voluto così tanto tempo prima che la psichiatria biologica sviluppasse test clinici e cosa fare al riguardo?Mol Psychiatry2012;17(12): 1174 1179. [PubMed]
7. Gaynes BN, Warden D, Trivedi MH, Wisniewski SR, Fava M, Rush JA. Cosa ci ha insegnato STAR * D? Risultati di una sperimentazione clinica pratica su larga scala per pazienti con depressionePsychiatr Serv. 2009;60(11): 1439 1445. [PubMed]
8. Fekadu A, Rane LJ, Wooderson SC, Markopoulou K, Poon L, Cleare AJ. Previsione del risultato a lungo termine della depressione resistente al trattamento nell'assistenza terziariaBr J Psychiatry. 2012;201(5): 369.[PubMed]
9. Fekadu A, Wooderson SC, Markopoulo K, Donaldson C, Papadopoulos A, Cleare AJ. Cosa succede ai pazienti con depressione resistente al trattamento? Una revisione sistematica degli studi sui risultati a medio e lungo termineJ Affect Disord. 2009;116(1 2): 4 11. [PubMed]
10. Trivedi M. Strategie di trattamento per migliorare e sostenere la remissione nel disturbo depressivo maggioreDialoghi Clin Neurosci. 2008;10(4): 377. [Articolo gratuito di PMC]ï ¿½[PubMed]
11. Fekadu A, Wooderson SC, Markopoulou K, Cleare AJ. Il metodo di stadiazione di Maudsley per la depressione resistente al trattamento: previsione del risultato a lungo termine e persistenza dei sintomiJ Clin Psychiatry. 2009;70(7): 952 957. [PubMed]
12. Bennabi D, Aouizerate B, El-Hage W, et al. Fattori di rischio per la resistenza al trattamento nella depressione unipolare: una revisione sistematicaJ Affect Disord. 2015;171: 137 141. [PubMed]
13. Serretti A, Olgiati P, Liebman MN, et al. Previsione clinica della risposta antidepressiva nei disturbi dell'umore: modelli multivariati lineari vs. modelli di rete neuralePsichiatria Res. 2007;152(2 3): 223 231.[PubMed]
14. Driessen E, Hollon SD. Terapia cognitivo comportamentale per disturbi dell'umore: efficacia, moderatori e mediatoriPsychiatr Clin North Am. 2010;33(3): 537 555. [Articolo gratuito di PMC]ï ¿½[PubMed]
15. Cleare A, Pariante C, Young A, et al. Membri del Consensus Meeting Linee guida basate sull'evidenza per il trattamento dei disturbi depressivi con antidepressivi: una revisione delle linee guida dell'associazione britannica del 2008 per la psicofarmacologia.J Psychopharmacol. 2015;29(5): 459 525. [PubMed]
16. Tunnard C, Rane LJ, Wooderson SC, et al. L'impatto delle avversità infantili sulla tendenza al suicidio e sul decorso clinico nella depressione resistente al trattamentoJ Affect Disord. 2014;152 154: 122 130. [PubMed]
17. Nemeroff CB, Heim CM, Thase ME, et al. Risposte differenziali alla psicoterapia rispetto alla farmacoterapia nei pazienti con forme croniche di depressione maggiore e traumi infantiliProc Natl Acad Sci US A. 2003;100(24): 14293 14296. [Articolo gratuito di PMC]ï ¿½[PubMed]
18. Nierenberg AA. Predittori di risposta agli antidepressivi principi generali e implicazioni clinichePsychiatr Clin North Am. 2003;26(2): 345 352. [PubMed]
19. Thase ME. Utilizzo di biomarcatori per prevedere la risposta al trattamento nel disturbo depressivo maggiore: prove da studi passati e presentiDialoghi Clin Neurosci. 2014;16(4): 539 544. [Articolo gratuito di PMC]ï ¿½[PubMed]
20. Jani BD, McLean G, Nicholl BI, et al. Valutazione del rischio e previsione dei risultati nei pazienti con sintomi depressivi: una revisione del potenziale ruolo dei biomarcatori basati sul sangue periferico.Front Hum Neurosci. 2015;9: 18. [Articolo gratuito di PMC]ï ¿½[PubMed]
21. Suravajhala P, Kogelman LJ, Kadarmideen HN. Integrazione e analisi multi-omiche dei dati utilizzando approcci di genomica dei sistemi: metodi e applicazioni nella produzione animale, salute e benessereGenet Sel Evol. 2016;48(1): 1. [Articolo gratuito di PMC]ï ¿½[PubMed]
22. Menke A. Espressione genica: biomarcatore della terapia antidepressiva? Psichiatria Int Rev. 2013;25(5): 579 591. [PubMed]
23. Peng B, Li H, Peng XX. Metabolomica funzionale: dalla scoperta di biomarcatori alla riprogrammazione del metabolomaCellula proteica2015;6(9): 628 637. [Articolo gratuito di PMC]ï ¿½[PubMed]
24. Aagaard K, Petrosino J, Keitel W, et al. La strategia del progetto Human Microbiome per un campionamento completo del microbioma umano e perché è importanteFASEB J. 2013;27(3): 1012.[Articolo gratuito di PMC]ï ¿½[PubMed]
25. Sonner Z, Wilder E, Heikenfeld J, et al. La microfluidica della ghiandola sudoripare eccrina, comprese le implicazioni di partizionamento, trasporto e biosensori dei biomarcatori.Biomicrofluidica . 2015;9(3): 031301.[Articolo gratuito di PMC]ï ¿½[PubMed]
26. Schmidt HD, Shelton RC, Duman RS. Biomarcatori funzionali della depressione: diagnosi, trattamento e fisiopatologiaNeuropsychopharm. 2011;36(12): 2375 2394. [Articolo gratuito di PMC]ï ¿½[PubMed]
27. J Brand S, Moller M, H Harvey B.Una revisione dei biomarcatori nei disturbi dell'umore e psicotici: una dissezione dei correlati clinici rispetto a quelli preclinici.Curr Neuropharmacol. 2015;13(3): 324.[Articolo gratuito di PMC]ï ¿½[PubMed]
28. Lopresti AL, Maker GL, Hood SD, Drummond PD. Una revisione dei biomarcatori periferici nella depressione maggiore: il potenziale dei biomarcatori dello stress infiammatorio e ossidativoProg Neuropsychopharmacol Biol Psychiatry2014;48: 102 111. [PubMed]
29. Fu CH, Steiner H, Costafreda SG. Biomarcatori neurali predittivi della risposta clinica nella depressione: una meta-analisi di studi di neuroimaging funzionale e strutturale di terapie farmacologiche e psicologiche.Neurobiol Dis. 2013;52: 75 83. [PubMed]
30. Mamdani F, Berlim M, Beaulieu M, Labbe A, Merette C, Turecki G. Biomarcatori di espressione genica della risposta al trattamento con citalopram nel disturbo depressivo maggiore.Transl Psychiatry. 2011;1(6): e13.[Articolo gratuito di PMC]ï ¿½[PubMed]
31. Smith RS. La teoria dei macrofagi della depressioneIpotesi Med . 1991;35(4): 298 306. [PubMed]
32. Irwin MR, Miller AH. Disturbi depressivi e immunità: 20 anni di progressi e scoperteBrain Behav Immun. 2007;21(4): 374 383. [PubMed]
33. Maes M, Leonard B, Myint A, Kubera M, Verkerk R. La nuova ipotesi di depressione 5-HT : l'attivazione immunitaria cellulo-mediata induce l'indoleamina 2,3-diossigenasi, che porta a un abbassamento del triptofano plasmatico e una maggiore sintesi di cataboliti dannosi del triptofano (TRYCAT), che contribuiscono entrambi all'insorgenza della depressione.Prog Neuropsychopharmacol Biol Psychiatry2011;35(3): 702.[PubMed]
34. Miller AH, Maletic V, Raison CL. Infiammazione e suo malcontento: il ruolo delle citochine nella fisiopatologia della depressione maggiore.Biol Psychiatry. 2009;65(9): 732 741. [Articolo gratuito di PMC]ï ¿½[PubMed]
35. Miller AH, Raison CL. Il ruolo dell'infiammazione nella depressione: dall'imperativo evolutivo al moderno target di trattamentoNat Rev Immun. 2016;16(1): 22 34. [Articolo gratuito di PMC]ï ¿½[PubMed]
36. Raison CL, Capuron L, Miller AH. Le citochine cantano il blues: infiammazione e patogenesi della depressioneTrends Immun. 2006;27(1): 24 31. [Articolo gratuito di PMC]ï ¿½[PubMed]
37. Raison CL, Felger JC, Miller AH. Infiammazione e resistenza al trattamento nella depressione maggiore: la tempesta perfettaPsychiatr Times. 2013;30(9)
38. Dowlati Y, Herrmann N, Swardfager W, et al. Una meta-analisi delle citochine nella depressione maggioreBiol Psychiatry. 2010;67(5): 446 457. [PubMed]
39. Eyre HA, Air T, Pradhan A, et al. Una meta-analisi delle chemochine nella depressione maggioreProg Neuropsychopharmacol Biol Psychiatry2016;68: 1 8. [Articolo gratuito di PMC]ï ¿½[PubMed]
40. Haapakoski R, Mathieu J, Ebmeier KP, Alenius H, Kivim ki M. Meta-analisi cumulativa delle interleuchine 6 e 1 ?, fattore di necrosi tumorale? e proteina C reattiva in pazienti con disturbo depressivo maggioreBrain Behav Immun. 2015;49: 206 215. [Articolo gratuito di PMC]ï ¿½[PubMed]
41. Howren MB, Lamkin DM, Suls J. Associazioni di depressione con proteina C-reattiva, IL-1 e IL-6: una meta-analisi.Psychosom Med. 2009;71(2): 171 186. [PubMed]
42. Liu Y, Ho RC-M, Mak A. L'interleuchina (IL) -6, il fattore di necrosi tumorale alfa (TNF-?) E i recettori solubili dell'interleuchina-2 (sIL-2R) sono elevati nei pazienti con disturbo depressivo maggiore: un meta- analisi e meta-regressioneJ Affect Disord. 2012;139(3): 230 239. [PubMed]
43. Strawbridge R, Arnone D, Danese A, Papadopoulos A, Herane Vives A, Cleare AJ. Infiammazione e risposta clinica al trattamento nella depressione: una meta-analisiEur Neuropsychopharmacol. 2015;25(10): 1532 1543. [PubMed]
44. Farooq RK, Asghar K, Kanwal S, Zulqernain A. Ruolo delle citochine infiammatorie nella depressione: concentrarsi sull'interleuchina-1? (Recensione) Biomed Rep. 2017;6(1): 15 20. [Articolo gratuito di PMC]ï ¿½[PubMed]
45. Cattaneo A, Ferrari C, Uher R, et al. Misurazioni assolute del fattore inibitorio della migrazione dei macrofagi e dell'interleuchina-1-? I livelli di mRNA predicono accuratamente la risposta al trattamento nei pazienti depressiInt J Neuropsychopharmacol. 2016;19(10): pyw045. [Articolo gratuito di PMC]ï ¿½[PubMed]
46. Baune B, Smith E, Reppermund S, et al. I biomarcatori infiammatori predicono sintomi depressivi, ma non ansia durante l'invecchiamento: lo studio prospettico sulla memoria e sull'invecchiamento di SydneyPsiconeuroendocrinolo . 2012;37(9): 1521 1530. [PubMed]
47. Fornaro M, Rocchi G, Escelsior A, Contini P, Martino M. I diversi trend delle citochine nei pazienti depressi che ricevono duloxetina indicano background biologici differenziali.J Affect Disord. 2013;145(3): 300 307. [PubMed]
48. Hernandez ME, Mendieta D, Martinez-Fong D, et al. Variazioni nei livelli di citochine circolanti durante il corso di 52 settimane di trattamento con SSRI per il disturbo depressivo maggioreEur Neuropsychopharmacol. 2008;18(12): 917 924. [PubMed]
49. Hannestad J, DellaGioia N, Bloch M. L'effetto del trattamento con farmaci antidepressivi sui livelli sierici di citochine infiammatorie: una meta-analisi.Neuropsicofarmacologia . 2011;36(12): 2452.[Articolo gratuito di PMC]ï ¿½[PubMed]
50. Hiles SA, Attia J, Baker AL. Cambiamenti dell'interleuchina-6, della proteina C-reattiva e dell'interleuchina-10 nelle persone con depressione in seguito a trattamento antidepressivo: una meta-analisi.Brain Behav Immun; Presentato a: 17th Meeting annuale della PsychoNeuroImmunology Research Society PsychoNeuroImmunology: Crossing Disciplines to Combat Disease; 2012. p. S44.
51. Harley J, Luty S, Carter J, Mulder R, Joyce P.Proteina C reattiva elevata nella depressione: un predittore di un buon esito a lungo termine con antidepressivi e scarso esito con la psicoterapia.J Psychopharmacol. 2010;24(4): 625 626. [PubMed]
52. Uher R, Tansey KE, Dew T, et al. Un biomarcatore infiammatorio come predittore differenziale dell'esito del trattamento della depressione con escitalopram e nortriptilina.Sono J Psychiatry. 2014;171(2): 1278.[PubMed]
53. Chang HH, Lee IH, Gean PW, et al. Risposta al trattamento e deterioramento cognitivo nella depressione maggiore: associazione con la proteina C reattivaBrain Behav Immun. 2012;26(1): 90 95. [PubMed]
54. Raison CL, Rutherford RE, Woolwine BJ, et al. Uno studio controllato randomizzato dell'antagonista del fattore di necrosi tumorale infliximab per la depressione resistente al trattamento: il ruolo dei biomarcatori infiammatori al basale.JAMA Psychiatry. 2013;70(1): 31 41. [Articolo gratuito di PMC]ï ¿½[PubMed]
55. Krishnadas R, Cavanagh J. Depression: an infiammatory disease? J Neurol Neurosurg Psychiatry. 2012;83(5): 495 502. [PubMed]
56. Raison CL, Miller AH. La depressione è un disturbo infiammatorio? Curr Psychiatry Rep. 2011;13(6): 467 475. [Articolo gratuito di PMC]ï ¿½[PubMed]
57. Simon N, McNamara K, Chow C, et al. Un esame dettagliato delle anomalie delle citochine nel Disturbo Depressivo MaggioreEur Neuropsychopharmacol. 2008;18(3): 230 233. [Articolo gratuito di PMC]ï ¿½[PubMed]
58. Dahl J, Ormstad H, Aass HC, et al. I livelli plasmatici di varie citochine sono aumentati durante la depressione in corso e vengono ridotti a livelli normali dopo il recuperoPsiconeuroendocrinolo . 2014;45: 77 86. [PubMed]
59. Stelzhammer V, Haenisch F, Chan MK, et al. Cambiamenti proteomici nel siero di pazienti affetti da depressione maggiore di prima insorgenza, na ve ai farmaci antidepressiviInt J Neuropsychopharmacol. 2014;17(10): 1599 1608. [PubMed]
60. Liu Y, HO RCM, Mak A. Il ruolo dell'interleuchina (IL) -17 nell'ansia e nella depressione dei pazienti con artrite reumatoide.Int J Rheum Dis. 2012;15(2): 183 187. [PubMed]
61. Diniz BS, Sibille E, Ding Y, et al. Biosignatura del plasma e patologia cerebrale correlata al deterioramento cognitivo persistente nella depressione in età avanzataMol Psychiatry2015;20(5): 594 601. [Articolo gratuito di PMC][PubMed]
62. Janelidze S, Ventorp F, Erhardt S, et al. Livelli alterati delle chemochine nel liquido cerebrospinale e nel plasma dei tentatori di suicidioPsiconeuroendocrinolo . 2013;38(6): 853 862. [PubMed]
63. Powell TR, Schalkwyk LC, Heffernan AL, et al. Il fattore di necrosi tumorale e i suoi bersagli nella via delle citochine infiammatorie sono identificati come biomarcatori trascrittomici putativi per la risposta all'escitalopram.Eur Neuropsychopharmacol. 2013;23(9): 1105 1114. [PubMed]
64. Wong M, Dong C, Maestre-Mesa J, Licinio J. I polimorfismi nei geni correlati all'infiammazione sono associati alla suscettibilità alla depressione maggiore e alla risposta antidepressiva.Mol Psychiatry2008;13(8): 800 812. [Articolo gratuito di PMC]ï ¿½[PubMed]
65. Kling MA, Alesci S, Csako G, et al. Stato proinfiammatorio di basso grado sostenuto in donne non medicate e rimesse con disturbo depressivo maggiore, come evidenziato da livelli sierici elevati delle proteine della fase acuta, proteina C-reattiva e amiloide sierica A.Biol Psychiatry. 2007;62(4): 309 313. [Articolo gratuito di PMC][PubMed]
66. Schaefer M, Sarkar S, Schwarz M, Friebe A. Molecola di adesione intracellulare solubile-1 in pazienti con disturbi affettivi unipolari o bipolari: risultati di uno studio pilota.Neuropsychobiol. 2016;74(1): 8.[PubMed]
67. Dimopoulos N, Piperi C, Salonicioti A, et al. Elevazione della concentrazione plasmatica delle molecole di adesione nella depressione in età avanzataInt J Geriatr Psychiatry. 2006;21(10): 965 971. [PubMed]
68. Bocchio-Chiavetto L, Bagnardi V, Zanardini R, et al. Livelli sierici e plasmatici di BDNF nella depressione maggiore: uno studio di replicazione e meta-analisiWorld J Biol Psychiatry2010;11(6): 763 773. [PubMed]
69. Brunoni AR, Lopes M, Fregni F.Una revisione sistematica e meta-analisi di studi clinici sulla depressione maggiore e sui livelli di BDNF: implicazioni per il ruolo della neuroplasticità nella depressione.Int J Neuropsychopharmacol. 2008;11(8): 1169 1180. [PubMed]
70. Molendijk M, Spinhoven P, Polak M, Bus B, Penninx B, Elzinga B. Concentrazioni sieriche di BDNF come manifestazioni periferiche di depressione: prove da una revisione sistematica e meta-analisi su 179 associazioni.Mol Psychiatry2014;19(7): 791 800. [PubMed]
71. Sen S, Duman R, Sanacora G. Fattore neurotrofico derivato dal cervello del siero, depressione e farmaci antidepressivi: meta-analisi e implicazioni.Biol Psychiatry. 2008;64(6): 527 532. [Articolo gratuito di PMC][PubMed]
72. Zhou L, Xiong J, Lim Y, et al. Upregulation del sangue proBDNF e dei suoi recettori nella depressione maggioreJ Affect Disord. 2013;150(3): 776 784. [PubMed]
73. Chen YW, Lin PY, Tu KY, Cheng YS, Wu CK, Tseng PT. Livelli di fattore di crescita nervoso significativamente più bassi nei pazienti con disturbo depressivo maggiore rispetto ai soggetti sani: una meta-analisi e una revisione sistematica.Neuropsychiatr Dis Treat. 2014;11: 925 933. [Articolo gratuito di PMC]ï ¿½[PubMed]
74. Lin PY, Tseng PT. Diminuzione dei livelli di fattori neurotrofici derivati dalla linea cellulare gliali in pazienti con depressione: uno studio meta-analitico.J Psychiatr Res. 2015;63: 20 27. [PubMed]
75. Warner-Schmidt JL, Duman RS. VEGF come potenziale bersaglio per l'intervento terapeutico nella depressioneCurr Op Pharmacol. 2008;8(1): 14 19. [Articolo gratuito di PMC]ï ¿½[PubMed]
76. Carvalho AF, K hler CA, McIntyre RS, et al. Fattore di crescita endoteliale vascolare periferico come nuovo biomarcatore della depressione: una meta-analisiPsiconeuroendocrinolo . 2015;62: 18 26. [PubMed]
77. Tseng PT, Cheng YS, Chen YW, Wu CK, Lin PY. Aumento dei livelli del fattore di crescita endoteliale vascolare nei pazienti con disturbo depressivo maggiore: una meta-analisiEur Neuropsychopharmacol. 2015;25(10): 1622 1630. [PubMed]
78. Carvalho L, Torre J, Papadopoulos A, et al. La mancanza di beneficio clinico terapeutico degli antidepressivi è associata all'attivazione complessiva del sistema infiammatorioJ Affect Disord. 2013;148(1): 136 140. [PubMed]
79. Clark-Raymond A, Meresh E, Hoppensteadt D, et al. Fattore di crescita endoteliale vascolare: potenziale predittore della risposta al trattamento nella depressione maggioreWorld J Biol Psychiatry2015: 1 11. [PubMed]
80. Isung J, Mobarrez F, Nordstr m P, sberg M, Jokinen J. Basso fattore di crescita dell'endotelio vascolare (VEGF) plasmatico associato al suicidio completato.World J Biol Psychiatry2012;13(6): 468 473. [PubMed]
81. Buttensch n HN, Foldager L, Elfving B, Poulsen PH, Uher R, Mors O. Fattori neurotrofici nella depressione in risposta al trattamento.J Affect Disord. 2015;183: 287 294. [PubMed]
82. Szcz? Sny E,? Lusarczyk J, G? Ombik K, et al. Possibile contributo dell'IGF-1 al disturbo depressivoPharmacol Rep. 2013;65(6): 1622 1631. [PubMed]
83. Tu KY, Wu MK, Chen YW, et al. Livelli di fattore di crescita insulino-simile periferico 1 significativamente più alti nei pazienti con disturbo depressivo maggiore o disturbo bipolare rispetto ai controlli sani: una meta-analisi e revisione secondo le linee guida di PRISMA.Med. 2016;95(4): e2411. [Articolo gratuito di PMC]ï ¿½[PubMed]
84. Wu CK, Tseng PT, Chen YW, Tu KY, Lin PY. Livelli di fattore di crescita 2 dei fibroblasti periferici significativamente più alti in pazienti con disturbo depressivo maggiore: una meta-analisi preliminare secondo le linee guida MOOSE.Med. 2016;95(33): e4563. [Articolo gratuito di PMC]ï ¿½[PubMed]
85. He S, Zhang T, Hong B, et al. Diminuzione dei livelli sierici del fattore di crescita 2 dei fibroblasti nei pazienti pre e post trattamento con disturbo depressivo maggiore.Neurosci Lett. 2014;579: 168 172. [PubMed]
86. Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Alterata espressione genica del fattore neurotrofico derivato dal cervello e del recettore tirosina chinasi B nel cervello post-mortem di soggetti suicidi.Arch Gen Psychiatry2003;60(8): 804 815. [PubMed]
87. Srikanthan K, Feyh A, Visweshwar H, Shapiro JI, Sodhi K.Revisione sistematica dei biomarcatori della sindrome metabolica: un pannello per la diagnosi precoce, la gestione e la stratificazione del rischio nella popolazione della Virginia occidentale.Int J Med Sci. 2016;13(1): 25. [Articolo gratuito di PMC]ï ¿½[PubMed]
88. Lu XY. L'ipotesi della leptina della depressione: un potenziale legame tra disturbi dell'umore e obesità? Curr Op Pharmacol. 2007;7(6): 648 652. [Articolo gratuito di PMC]ï ¿½[PubMed]
89. Wittekind DA, Kluge M. Ghrelin nei disturbi psichiatrici (una revisione)Psiconeuroendocrinolo . 2015;52: 176 194. [PubMed]
90. Kan C, Silva N, Golden SH, et al. Una revisione sistematica e una meta-analisi dell'associazione tra depressione e resistenza all'insulinaCura del diabete2013;36(2): 480 489. [Articolo gratuito di PMC]ï ¿½[PubMed]
91. Liu X, Li J, Zheng P, et al. La lipidomica del plasma rivela potenziali marcatori lipidici del disturbo depressivo maggioreAnal Bioanal Chem. 2016;408(23): 6497 6507. [PubMed]
92. Lustman PJ, Anderson RJ, Freedland KE, De Groot M, Carney RM, Clouse RE. Depressione e scarso controllo glicemico: una revisione meta-analitica della letteraturaCura del diabete2000;23(7): 934 942. [PubMed]
93. Maes M. Evidenza per una risposta immunitaria nella depressione maggiore: una revisione e un'ipotesiProg NeuroPsychopharmacol Biol Psychiatry1995;19(1): 11 38. [PubMed]
94. Zheng H, Zheng P, Zhao L, et al. Diagnosi predittiva della depressione maggiore utilizzando la metabolomica basata su NMR e la macchina vettoriale di supporto dei minimi quadratiClinica Chimica Acta. 2017;464: 223 227.[PubMed]
95. Xia Q, Wang G, Wang H, Xie Z, Fang Y, Li Y. Studio del metabolismo del glucosio e dei lipidi in pazienti con depressione del primo episodio.J Clin Psychiatry. 2009;19: 241 243.
96. Kaufman J, DeLorenzo C, Choudhury S, Parsey RV. Il recettore 5-HT 1A nel disturbo depressivo maggioreEur Neuropsychopharmacology. 2016;26(3): 397 410. [Articolo gratuito di PMC]ï ¿½[PubMed]
97. Jacobsen JP, Krystal AD, Krishnan KRR, Caron MG. 5-idrossitriptofano aggiuntivo a rilascio lento per la depressione resistente al trattamento: razionale clinico e preclinico.Tendenze Pharmacol Sci. 2016;37(11): 933 944. [Articolo gratuito di PMC]ï ¿½[PubMed]
98. Salamone JD, Correa M, Yohn S, Cruz LL, San Miguel N, Alatorre L. La farmacologia del comportamento di scelta correlato allo sforzo: dopamina, depressione e differenze individuali.Processi di comportamento2016;127: 3 17. [PubMed]
99. Coplan JD, Gopinath S, Abdallah CG, Berry BR. Un'ipotesi neurobiologica di depressione resistente al trattamento meccanismi per la non efficacia degli inibitori selettivi della ricaptazione della serotonina.Front Behav Neurosci. 2014;8: 189. [Articolo gratuito di PMC]ï ¿½[PubMed]
100. Popa D, Cerdan J, Reprant C, et al. Uno studio longitudinale sul deflusso di 5-HT durante il trattamento con fluoxetina cronica utilizzando una nuova tecnica di microdialisi cronica in un ceppo di topo altamente emotivo.Eur J Pharmacol. 2010;628(1): 83 90. [PubMed]
101. Atake K, Yoshimura R, Hori H, et al. La duloxetina, un inibitore selettivo della ricaptazione della noradrenalina, ha aumentato i livelli plasmatici del 3-metossi-4-idrossifenilglicole ma non dell'acido omovanillico nei pazienti con disturbo depressivo maggiore.Clin Psychopharmacol Neurosci. 2014;12(1): 37 40. [Articolo gratuito di PMC][PubMed]
102. Ueda N, Yoshimura R, Shinkai K, Nakamura J. I livelli plasmatici dei metaboliti delle catecolamine predicono la risposta alla sulpiride o alla fluvoxamina nella depressione maggiore.Farmacopsichiatria . 2002;35(05): 175.[PubMed]
103. Yamana M, Atake K, Katsuki A, Hori H, Yoshimura R. Marcatori biologici del sangue per la previsione della risposta di escitalopram in pazienti con disturbo depressivo maggiore: studio preliminare.J Deprimere l'ansia2016;5: 222.
104. Parker KJ, Schatzberg AF, Lyons DM. Aspetti neuroendocrini dell'ipercortisolismo nella depressione maggioreHorm Behav. 2003;43(1): 60 66. [PubMed]
105. Stetler C, Miller GE. Depressione e attivazione ipotalamo-ipofisi-surrenalica: una sintesi quantitativa di quattro decenni di ricercaPsychosom Med. 2011;73(2): 114 126. [PubMed]
106. Herane Vives A, De Angel V, Papadopoulos A, et al. La relazione tra cortisolo, stress e malattie psichiatriche: nuove intuizioni utilizzando l'analisi dei capelliJ Psychiatr Res. 2015;70: 38 49. [PubMed]
107. Fischer S, Strawbridge R, Vives AH, Cleare AJ. Cortisolo come predittore della risposta alla terapia psicologica nei disturbi depressivi: revisione sistematica e meta-analisiBr J Psychiatry. 2017;210(2): 105 109. [PubMed]
108. Anacker C, Zunszain PA, Carvalho LA, Pariante CM. Il recettore dei glucocorticoidi: perno della depressione e del trattamento antidepressivo? Psiconeuroendocrinologia . 2011;36(3): 415 425. [Articolo gratuito di PMC][PubMed]
109. Markopoulou K, Papadopoulos A, Juruena MF, Poon L, Pariante CM, Cleare AJ. Il rapporto cortisolo / DHEA nella depressione resistente al trattamentoPsiconeuroendocrinolo . 2009;34(1): 19 26. [PubMed]
110. Joffe RT, Pearce EN, Hennessey JV, Ryan JJ, Stern RA. Ipotiroidismo subclinico, umore e cognizione negli anziani: una revisioneInt J Geriatr Psychiatry. 2013;28(2): 111 118. [Articolo gratuito di PMC][PubMed]
111. Duval F, Mokrani MC, Erb A, et al. Stato cronobiologico dell'asse ipotalamico-ipofisario-tiroideo ed esito antidepressivo nella depressione maggiore.Psiconeuroendocrinolo . 2015;59: 71 80. [PubMed]
112. Marsden W. Plasticità sinaptica nella depressione: correlati molecolari, cellulari e funzionaliProg Neuropsychopharmacol Biol Psychiatry2013;43: 168 184. [PubMed]
113. Duman RS, Voleti B. Vie di segnalazione alla base della fisiopatologia e del trattamento della depressione: nuovi meccanismi per agenti ad azione rapida.Trends Neurosci. 2012;35(1): 47.[Articolo gratuito di PMC]ï ¿½[PubMed]
114. Ripke S, Wray NR, Lewis CM, et al. Una mega-analisi di studi di associazione su tutto il genoma per il disturbo depressivo maggioreMol Psychiatry2013;18(4): 497 511. [Articolo gratuito di PMC]ï ¿½[PubMed]
115. Mullins N, Power R, Fisher H, et al. Interazioni poligeniche con avversità ambientali nell'eziologia del disturbo depressivo maggiorePsychol Med. 2016;46(04): 759 770. [Articolo gratuito di PMC]ï ¿½[PubMed]
116. Lewis S. Disturbi neurologici: telomeri e depressioneNat Rev Neurosci. 2014;15(10): 632.[PubMed]
117. Lindqvist D, Epel ES, Mellon SH, et al. Disturbi psichiatrici e lunghezza dei telomeri dei leucociti: meccanismi sottostanti che collegano la malattia mentale con l'invecchiamento cellulare.Neurosci Biobehav Rev. 2015;55: 333 364. [Articolo gratuito di PMC]ï ¿½[PubMed]
118. McCall WV. Un biomarcatore dell'attività a riposo per prevedere la risposta agli SSRI nel disturbo depressivo maggioreJ Psychiatr Res. 2015;64: 19 22. [Articolo gratuito di PMC]ï ¿½[PubMed]
119. Schuch FB, Deslandes AC, Stubbs B, Gosmann NP, da Silva CTB, de Almeida Fleck MP. Effetti neurobiologici dell'esercizio sul disturbo depressivo maggiore: una revisione sistematicaNeurosci Biobehav Rev. 2016;61: 1 11. [PubMed]
120. Foster JA, Neufeld K-AM. Asse intestinale: come il microbioma influenza l'ansia e la depressioneTrends Neurosci. 2013;36(5): 305 312. [PubMed]
121. Quattrocki E, Baird A, Yurgelun-Todd D. Aspetti biologici del legame tra fumo e depressione.Psichiatria di Harv Rev. 2000;8(3): 99 110. [PubMed]
122. Maes M, Kubera M, Obuchowiczwa E, Goehler L, Brzeszcz J. Depression molteplici comorbidità spiegate da percorsi di stress (neuro) infiammatorio e ossidativo e nitrosativo.Neuro Endocrinol Lett. 2011;32(1): 7 24. [PubMed]
123. Miller G, Rohleder N, Cole SW. Lo stress interpersonale cronico predice l'attivazione di vie di segnalazione pro e antinfiammatorie sei mesi dopoPsychosom Med. 2009;71(1): 57. [Articolo gratuito di PMC][PubMed]
124. Steptoe A, Hamer M, Chida Y. Gli effetti dello stress psicologico acuto sui fattori infiammatori circolanti negli esseri umani: una revisione e una meta-analisi.Brain Behav Immun. 2007;21(7): 901 912. [PubMed]
125. Danese A, Moffitt TE, Harrington H, et al. Esperienze infantili avverse e fattori di rischio degli adulti per malattie legate all'età: depressione, infiammazione e raggruppamento dei marcatori di rischio metabolico.Arch Pediatr Adolesc Med. 2009;163(12): 1135 1143. [Articolo gratuito di PMC]ï ¿½[PubMed]
126. Danese A, Pariante CM, Caspi A, Taylor A, Poulton R. Il maltrattamento infantile predice l'infiammazione degli adulti in uno studio sul corso della vita.Proc Natl Acad Sci US A. 2007;104(4): 1319 1324. [Articolo gratuito di PMC][PubMed]
127. Danese A, Caspi A, Williams B, et al. Incorporamento biologico dello stress attraverso processi infiammatori nell'infanziaMol Psychiatry2011;16(3): 244 246. [Articolo gratuito di PMC]ï ¿½[PubMed]
128. Suzuki A, Poon L, Kumari V, Cleare AJ. I pregiudizi di paura nell'elaborazione del volto emotivo a seguito di un trauma infantile come indicatore di resilienza e vulnerabilità alla depressioneChild Maltreat2015;20(4): 240 250. [PubMed]
129. Strawbridge R, Young AH. Asse HPA e disregolazione cognitiva nei disturbi dell'umore. In: McIntyre RS, Cha DS, editoriCompromissione cognitiva nel disturbo depressivo maggiore: rilevanza clinica, substrati biologici e opportunità di trattamento. Cambridge: Cambridge University Press; 2016. pagg. 179.
130. Keller J, Gomez R, Williams G, et al. Asse HPA nella depressione maggiore: cortisolo, sintomatologia clinica e variazione genetica predicono la cognizioneMol Psychiatry2016 agosto 16; Epub. [Articolo gratuito di PMC]ï ¿½[PubMed]
131. Hanson ND, Owens MJ, Nemeroff CB. Depressione, antidepressivi e neurogenesi: una rivalutazione criticaNeuropsychopharmacol. 2011;36(13): 2589 2602. [Articolo gratuito di PMC]ï ¿½[PubMed]
132. Chen Y, Baram TZ. Verso la comprensione di come lo stress nei primi anni di vita riprogramma le reti cerebrali cognitive ed emotiveNeuropsychopharmacol. 2015;41(1): 197 206. [Articolo gratuito di PMC]ï ¿½[PubMed]
133. Porter RJ, Gallagher P, Thompson JM, Young AH. Compromissione neurocognitiva in pazienti senza farmaci con disturbo depressivo maggioreBr J Psychiatry. 2003;182: 214 220. [PubMed]
134. Gallagher P, Robinson L, Gray J, Young A, Porter R. Funzione neurocognitiva a seguito di remissione nel disturbo depressivo maggiore: potenziale marker obiettivo di risposta? Aust NZJ Psychiatry2007;41(1): 54 61. [PubMed]
135. Pittenger C, Duman RS. Stress, depressione e neuroplasticità: una convergenza di meccanismiNeuropsychopharmacol. 2008;33(1): 88 109. [PubMed]
136. B ckman L, Nyberg L, Lindenberger U, Li SC, Farde L. La triade correlativa tra invecchiamento, dopamina e cognizione: stato attuale e prospettive future.Neurosci Biobehav Rev. 2006;30(6): 791 807. [PubMed]
137. Allison DJ, Ditor DS. L'eziologia infiammatoria comune della depressione e del deterioramento cognitivo: un obiettivo terapeuticoJ Neuroinfiammazione2014;11: 151. [Articolo gratuito di PMC]ï ¿½[PubMed]
138. Rosenblat JD, Brietzke E, Mansur RB, Maruschak NA, Lee Y, McIntyre RS. L'infiammazione come substrato neurobiologico del deterioramento cognitivo nel disturbo bipolare: prove, fisiopatologia e implicazioni del trattamento.J Affect Disord. 2015;188: 149 159. [PubMed]
139. Krogh J, Benros ME, Jrgensen MB, Vesterager L, Elfving B, Nordentoft M. L'associazione tra sintomi depressivi, funzione cognitiva e infiammazione nella depressione maggiore.Brain Behav Immun. 2014;35: 70 76. [PubMed]
140. Soares CN, Zitek B. Sensibilità agli ormoni riproduttivi e rischio di depressione durante il ciclo di vita femminile: un continuum di vulnerabilità? J Psychiatry Neurosci. 2008;33(4): 331. [Articolo gratuito di PMC]ï ¿½[PubMed]
141. Hiles SA, Baker AL, de Malmanche T, Attia J. Una meta-analisi delle differenze in IL-6 e IL-10 tra persone con e senza depressione: esplorare le cause dell'eterogeneità.Brain Behav Immun. 2012;26(7): 1180 1188. [PubMed]
142. Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. La secrezione di adipochine grasse viscerali è associata all'infiammazione sistemica negli esseri umani obesi.Diabete2007;56(4): 1010 1013. [PubMed]
143. Divani AA, Luo X, Datta YH, Flaherty JD, Panoskaltsis-Mortari A. Effetto dei contraccettivi ormonali orali e vaginali sui biomarcatori del sangue infiammatorio.Mediatori Inflamm. 2015;2015: 379501.[Articolo gratuito di PMC]ï ¿½[PubMed]
144. Ramsey JM, Cooper JD, Penninx BW, Bahn S. Variazione nei biomarcatori sierici con sesso e stato ormonale femminile: implicazioni per i test clinici.Sci Rep. 2016;6: 26947. [Articolo gratuito di PMC]ï ¿½[PubMed]
145. Eyre H, Lavretsky H, Kartika J, Qassim A, Baune B.Effetti modulatori delle classi di antidepressivi sul sistema immunitario innato e adattativo nella depressione.Farmacopsichiatria . 2016;49(3): 85.[Articolo gratuito di PMC]ï ¿½[PubMed]
146. Hiles SA, Baker AL, de Malmanche T, Attia J. Interleukin-6, proteina C-reattiva e interleuchina-10 dopo il trattamento antidepressivo nelle persone con depressione: una meta-analisi.Psychol Med. 2012;42(10): 2015 2026. [PubMed]
147. Janssen DG, Caniato RN, Verster JC, Baune BT. Una revisione psiconeuroimmunologica sulle citochine coinvolte nella risposta al trattamento antidepressivoHum Psychopharmacol. 2010;25(3): 201 215. [PubMed]
148. Artigas F. Recettori della serotonina coinvolti negli effetti antidepressiviPharmacol Ther. 2013;137(1): 119 131. [PubMed]
149. Lee BH, Kim YK. I ruoli del BDNF nella fisiopatologia della depressione maggiore e nel trattamento antidepressivoIndagine psichiatrica2010;7(4): 231 235. [Articolo gratuito di PMC]ï ¿½[PubMed]
150. Hashimoto K. Biomarcatori infiammatori come predittori differenziali della risposta antidepressivaInt J Mol Sci. 2015;16(4): 7796 7801. [Articolo gratuito di PMC]ï ¿½[PubMed]
151. Goldberg D. L'eterogeneità della "depressione maggiore"Psichiatria mondiale2011;10(3): 226.[Articolo gratuito di PMC]ï ¿½[PubMed]
152. Arnow BA, Blasey C, Williams LM, et al. Sottotipi di depressione nella previsione della risposta antidepressiva: un rapporto dallo studio iSPOT-DSono J Psychiatry. 2015;172(8): 743 750. [PubMed]
153. Kunugi H, Hori H, Ogawa S. Marcatori biochimici che sottotipizzano il disturbo depressivo maggiorePsychiatry Clin Neurosci. 2015;69(10): 597 608. [PubMed]
154. Baune B, Stuart M, Gilmour A, et al. La relazione tra sottotipi di depressione e malattie cardiovascolari: una revisione sistematica dei modelli biologiciTransl Psychiatry. 2012;2(3): e92.[Articolo gratuito di PMC]ï ¿½[PubMed]
155. Vogelzangs N, Duivis HE, Beekman AT, et al. Associazione di disturbi depressivi, caratteristiche della depressione e farmaci antidepressivi con l'infiammazioneTransl Psychiatry. 2012;2: e79.[Articolo gratuito di PMC]ï ¿½[PubMed]
156. Lamers F, Vogelzangs N, Merikangas K, De Jonge P, Beekman A, Penninx B.Evidenza di un ruolo differenziale della funzione dell'asse HPA, dell'infiammazione e della sindrome metabolica nella depressione malinconica rispetto a quella atipica.Mol Psychiatry2013;18(6): 692 699. [PubMed]
157. Penninx BW, Milaneschi Y, Lamers F, Vogelzangs N. Capire le conseguenze somatiche della depressione: meccanismi biologici e ruolo del profilo dei sintomi depressivi.BMC Med. 2013;11(1): 1.[Articolo gratuito di PMC]ï ¿½[PubMed]
158. Capuron L, Su S, Miller AH, et al. Sintomi depressivi e sindrome metabolica: l'infiammazione è l'anello sottostante? Biol Psychiatry. 2008;64(10): 896 900. [Articolo gratuito di PMC]ï ¿½[PubMed]
159. Dantzer R, O'Connor JC, Freund GG, Johnson RW, Kelley KW. Dall'infiammazione alla malattia e alla depressione: quando il sistema immunitario soggioga il cervelloNat Rev Neurosci. 2008;9(1): 46.[Articolo gratuito di PMC]ï ¿½[PubMed]
160. Maes M, Berk M, Goehler L, et al. La depressione e il comportamento di malattia sono risposte affrontate da Janus a percorsi infiammatori condivisiBMC Med. 2012;10: 66. [Articolo gratuito di PMC]ï ¿½[PubMed]
161. Merikangas KR, Jin R, He JP, et al. Prevalenza e correlati del disturbo dello spettro bipolare nell'iniziativa di indagine sulla salute mentale mondialeArch Gen Psychiatry2011;68(3): 241 251. [Articolo gratuito di PMC][PubMed]
162. Hirschfeld RM, Lewis L, Vornik LA. Percezioni e impatto del disturbo bipolare: fino a che punto siamo davvero arrivati? Risultati dell'indagine 2000 dell'associazione nazionale depressiva e maniaco-depressiva sugli individui con disturbo bipolareJ Clin Psychiatry. 2003;64(2): 161 174. [PubMed]
163. Young AH, MacPherson H. Rilevazione del disturbo bipolareBr J Psychiatry. 2011;199(1): 3.[PubMed]
164. V hringer PA, Perlis RH. Discriminare tra disturbo bipolare e disturbo depressivo maggiorePsychiatr Clin North Am. 2016;39(1): 1 10. [PubMed]
165. Becking K, Spijker AT, Hoencamp E, Penninx BW, Schoevers RA, Boschloo L. Disturbi dell'asse ipotalamo-ipofisi-surrene e attività immunologica che differenziano tra episodi depressivi unipolari e bipolari.PLoS One. 2015;10(7): e0133898. [Articolo gratuito di PMC]ï ¿½[PubMed]
166. Huang TL, Lin FC. Livelli di proteina C reattiva ad alta sensibilità in pazienti con disturbo depressivo maggiore e mania bipolareProg NeuroPsychopharmacol Biol Psychiatry2007;31(2): 370 372. [PubMed]
167. Angst J, Gamma A, Endrass J. Fattori di rischio per gli spettri bipolari e depressiviActa Psychiatr Scand. 2003;418: 15 19. [PubMed]
168. Fekadu A, Wooderson S, Donaldson C, et al. Uno strumento multidimensionale per quantificare la resistenza al trattamento nella depressione: il metodo di stadiazione di MaudsleyJ Clin Psychiatry. 2009;70(2): 177. [PubMed]
169. Papakostas G, Shelton R, Kinrys G, et al. Valutazione di un test diagnostico biologico basato su siero multiplo per il disturbo depressivo maggiore: uno studio pilota e di replicazione.Mol Psychiatry2013;18(3): 332 339. [PubMed]
170. Fan J, Han F, Liu H. Sfide dell'analisi dei big dataNatl Sci Rev. 2014;1(2): 293.[Articolo gratuito di PMC]ï ¿½[PubMed]
171. Li L, Jiang H, Qiu Y, Ching WK, Vassiliadis VS. Scoperta dei biomarcatori dei metaboliti: analisi del flusso e approccio alla rete di reazione-reazioneBMC Syst Biol. 2013;7(Suppl 2): S13. [Articolo gratuito di PMC][PubMed]
172. Patel MJ, Khalaf A, Aizenstein HJ. Studiare la depressione utilizzando metodi di imaging e apprendimento automaticoNeuroImage Clin. 2016;10: 115 123. [Articolo gratuito di PMC]ï ¿½[PubMed]
173. Lanquillon S, Krieg JC, Bening-Abu-Shach U, Vedder H. Produzione di citochine e risposta al trattamento nel disturbo depressivo maggiore.Neuropsychopharmacol. 2000;22(4): 370 379. [PubMed]
174. Lindqvist D, Janelidze S, Erhardt S, Tr skman-Bendz L, Engstr m G, Brundin L. CSF biomarcatori in tentters suicide una componente principale di analisi.Acta Psychiatr Scand. 2011;124(1): 52 61. [PubMed]
175. Hidalgo-Mazzei D, Murru A, Reinares M, Vieta E, Colom F. Big data in mental health: a sfidante frammentato futuro.Psichiatria mondiale2016;15(2): 186 187. [Articolo gratuito di PMC]ï ¿½[PubMed]
176. Consorzio C-DGotPG Identificazione di loci di rischio con effetti condivisi su cinque principali disturbi psichiatrici: un'analisi dell'intero genoma.Lancet. 2013;381(9875): 1371 1379. [Articolo gratuito di PMC]ï ¿½[PubMed]
177. Dipnall JF, Pasco JA, Berk M, et al. Fondere data mining, apprendimento automatico e statistiche tradizionali per rilevare biomarcatori associati alla depressionePLoS One. 2016;11(2): e0148195. [Articolo gratuito di PMC][PubMed]
178. K hler O, Benros ME, Nordentoft M, et al. Effetto del trattamento antinfiammatorio su depressione, sintomi depressivi ed effetti avversi: una revisione sistematica e una meta-analisi di studi clinici randomizzati.JAMA Psychiatry. 2014;71(12): 1381 1391. [PubMed]
179. Wolkowitz OM, Reus VI, Chan T, et al. Trattamento antiglucocorticoide della depressione: ketoconazolo in doppio ciecoBiol Psychiatry. 1999;45(8): 1070 1074. [PubMed]
180. McAllister-Williams RH, Anderson IM, Finkelmeyer A, et al. Aumento degli antidepressivi con metirapone per la depressione resistente al trattamento (studio ADD): uno studio in doppio cieco, randomizzato, controllato con placebo.Lancet Psychiatry2016;3(2): 117 127. [PubMed]
181. Gallagher P, Young AH. Trattamento con mifepristone (RU-486) per la depressione e la psicosi: una revisione delle implicazioni terapeuticheNeuropsychiatr Dis Treat. 2006;2(1): 33 42. [Articolo gratuito di PMC]ï ¿½[PubMed]
182. Otte C, Hinkelmann K, Moritz S, et al. Modulazione del recettore dei mineralcorticoidi come trattamento aggiuntivo nella depressione: uno studio proof-of-concept randomizzato, in doppio cieco, controllato con placebo.J Psychiatr Res. 2010;44(6): 339 346. [PubMed]
183. Ozbolt LB, Nemeroff CB. Modulazione dell'asse HPA nel trattamento dei disturbi dell'umoreDisturbo psichiatrico2013;51: 1147 1154.
184. Walker AK, Budac DP, Bisulco S, et al. Il blocco del recettore NMDA da parte della ketamina abroga il comportamento depressivo indotto dai lipopolisaccaridi nei topi C57BL / 6J.Neuropsychopharmacol. 2013;38(9): 1609 1616. [Articolo gratuito di PMC]ï ¿½[PubMed]
185. Lespérance F, Frasure-Smith N, St-André E, Turecki G, Lespérance P, Wisniewski SR. L'efficacia della supplementazione di omega-3 per la depressione maggiore: uno studio controllato randomizzatoJ Clin Psychiatry. 2010;72(8): 1054 1062. [PubMed]
186. Kim S, Bae K, Kim J, et al. L'uso di statine per il trattamento della depressione in pazienti con sindrome coronarica acutaTransl Psychiatry. 2015;5(8): e620. [Articolo gratuito di PMC]ï ¿½[PubMed]
187. Shishehbor MH, Brennan ML, Aviles RJ, et al. Le statine promuovono potenti effetti antiossidanti sistemici attraverso specifiche vie infiammatorieCircolazione2003;108(4): 426 431. [PubMed]
188. Mercier A, Auger-Aubin I, Lebeau JP, et al. Evidenza della prescrizione di antidepressivi per condizioni non psichiatriche nelle cure primarie: un'analisi delle linee guida e revisioni sistematiche.BMC Family Practice. 2013;14(1): 55. [Articolo gratuito di PMC]ï ¿½[PubMed]
189. Freland L, Beaulieu JM. Inibizione di GSK3 da parte del litio, da singole molecole a reti di segnalazioneFront Mol Neurosci. 2012;5: 14. [Articolo gratuito di PMC]ï ¿½[PubMed]
190. Horowitz MA, Zunszain PA. Anomalie neuroimmuni e neuroendocrine nella depressione: due facce della stessa medagliaAnn NY Acad Sci. 2015;1351(1): 68 79. [PubMed]
191. Juruena MF, Cleare AJ. Sovrapposizione tra depressione atipica, disturbo affettivo stagionale e sindrome da stanchezza cronicaRev Bras Psiquiatr. 2007;29: S19 S26. [PubMed]
192. Castr n E, Kojima M. Fattore neurotrofico derivato dal cervello nei disturbi dell'umore e nei trattamenti antidepressivi.Neurobiol Dis. 2017;97(Parte B): 119 126. [PubMed]
193. Pan A, Keum N, Okereke OI, et al. Associazione bidirezionale tra depressione e sindrome metabolica una revisione sistematica e una meta-analisi di studi epidemiologiciCura del diabete2012;35(5): 1171 1180. [Articolo gratuito di PMC]ï ¿½[PubMed]
194. Carvalho AF, Rocha DQ, McIntyre RS, et al. Le adipochine come biomarcatori emergenti della depressione: una revisione sistematica e una meta-analisiJ Psychiatric Res. 2014;59: 28 37. [PubMed]
195. Wise T, Cleare AJ, Herane A, Young AH, Arnone D.Utilità diagnostica e terapeutica del neuroimaging nella depressione: una panoramica.Neuropsychiatr Dis Treat. 2014;10: 1509 1522.[Articolo gratuito di PMC]ï ¿½[PubMed]
196. Tamatam A, Khanum F, Bawa AS. Biomarcatori genetici della depressioneIndian J Hum Genet2012;18(1): 20. [Articolo gratuito di PMC]ï ¿½[PubMed]
197. Yoshimura R, Nakamura J, Shinkai K, Ueda N. Risposta clinica al trattamento antidepressivo e livelli di 3-metossi-4-idrossifenilglicole: mini revisione.Prog Neuropsychopharmacol Biol Psychiatry2004;28(4): 611 616. [PubMed]
198. Pierscionek T, Adekunte O, Watson S, Ferrier N, Alabi A. Ruolo dei corticosteroidi nella risposta antidepressiva.ChronoPhys Ther. 2014;4: 87 98.
199. Hage MP, Azar ST. Il legame tra funzione tiroidea e depressioneRicerca tiroide J. 2012;2012: 590648. [Articolo gratuito di PMC]ï ¿½[PubMed]
200. Dunn EC, Brown RC, Dai Y, et al. Determinanti genetici della depressione: scoperte recenti e direzioni futurePsichiatria di Harv Rev. 2015;23(1): 1. [Articolo gratuito di PMC]ï ¿½[PubMed]
201. Yang CC, Hsu YL. Una revisione dei rilevatori di movimento indossabili basati sull'accelerometria per il monitoraggio dell'attività fisicaSensori2010;10(8): 7772 7788. [Articolo gratuito di PMC]ï ¿½[PubMed]
Artrite il dolore è un fenomeno complesso che coinvolge un'elaborazione neurofisiologica intricata a tutti i livelli del percorso del dolore. Le opzioni di trattamento disponibili per alleviare il dolore articolare sono piuttosto limitate e la maggior parte dei pazienti affetti da artrite riporta solo un modesto sollievo dal dolore con i trattamenti attuali. Una migliore comprensione dei meccanismi neurali responsabili del dolore muscoloscheletrico e l'identificazione di nuovi bersagli aiuterà a sviluppare future terapie farmacologiche. Questo articolo esamina alcune delle ultime ricerche sui fattori che contribuiscono al dolore articolare e copre aree come i cannabinoidi, i recettori attivati dalla proteinasi, i canali del sodio, le citochine e i potenziali canali dei recettori transitori. Viene anche discussa l'ipotesi emergente che l'osteoartrosi possa avere una componente neuropatica.
Introduzione
L'organizzazione mondiale della sanità classifica i disturbi muscoloscheletrici come la causa più frequente di disabilità nel mondo moderno, colpendo uno su tre adulti [1]. Ancora più allarmante è che la prevalenza di queste malattie è in aumento, mentre la nostra conoscenza delle cause sottostanti è piuttosto rudimentale.
Fig. 1 Uno schema che illustra alcuni dei target noti per modulare il dolore articolare. I neuromodulatori possono essere rilasciati dai terminali nervosi, dai mastociti e dai macrofagi per alterare la meccanosensibilità afferente. Gli endovanilloidi, l'acido e il calore nocivo possono attivare i canali ionici vanilloidi di tipo 1 (TRPV1) del recettore transitorio che portano al rilascio della sostanza algogena P (SP), che successivamente si lega ai recettori della neurochinina-1 (NK1). Le proteasi possono scindere e stimolare i recettori attivati dalla proteasi (PAR). Finora, PAR2 e PAR4 hanno dimostrato di sensibilizzare le afferenze primarie articolari. L'endocannabinoide anandamide (AE) è prodotto su richiesta e sintetizzato dalla N-arachidonoil fosfatidiletanolammina (NAPE) sotto l'azione enzimatica delle fosfolipasi. Una porzione di AE si lega quindi ai recettori dei cannabinoidi-1 (CB1) portando alla desensibilizzazione neuronale. L'AE non legato viene rapidamente assorbito da un trasportatore di membrana dell'anandamide (AMT) prima di essere scomposto da un'ammide idrolasi dell'acido grasso (FAAH) in etanolammina (Et) e acido arachidonico (AA). Le citochine fattore di necrosi tumorale-?(TNF-?), interleuchina-6 (IL-6) e interleuchina1-beta (IL-1?) possono legarsi ai rispettivi recettori per aumentare la trasmissione del dolore. Infine, i canali del sodio resistenti alla tetrodotossina (TTX) (Nav1.8) sono coinvolti nella sensibilizzazione neuronale.
I pazienti desiderano il loro il dolore cronico scomparire; tuttavia, gli analgesici attualmente prescritti sono in gran parte inefficaci e sono accompagnati da un'ampia gamma di effetti collaterali indesiderati. Pertanto, milioni di persone in tutto il mondo soffrono degli effetti debilitanti del dolore articolare, per i quali non esiste un trattamento soddisfacente [2].
Più di 100 diverse forme di artrite hanno l'osteoartrite (OA) come la più comune. L'OA è una malattia articolare progressivamente degenerativa che causa dolore cronico e perdita di funzionalità. Comunemente, l'OA è l'incapacità dell'articolazione di riparare efficacemente i danni in risposta a forze eccessive che vengono applicate su di essa. I fattori biologici e psicosociali che compongono il dolore cronico dell'OA non sono ben compresi, sebbene la ricerca in corso sveli la natura complessa dei sintomi della malattia [2]. Le attuali terapie, come i farmaci antinfiammatori non steroidei (FANS), forniscono un sollievo sintomatico, riducendo il dolore per brevi periodi di tempo, ma non alleviano il dolore per tutta la durata della vita del paziente. Inoltre, i FANS ad alte dosi non possono essere assunti ripetutamente per molti anni, poiché ciò può portare a tossicità renale e sanguinamento gastrointestinale.
Tradizionalmente, la ricerca sull'artrite si è concentrata in gran parte sulla cartilagine articolare come obiettivo primario per lo sviluppo terapeutico di nuovi farmaci per l'osteoartrite per la modificazione della malattia. Questa attenzione condrogenica ha gettato nuova luce sugli intricati fattori biochimici e biomeccanici che influenzano il comportamento dei condrociti nelle articolazioni malate. Tuttavia, poiché la cartilagine articolare è aneurale e avascolare, è improbabile che questo tessuto sia la fonte del dolore da OA. Questo fatto, unito ai risultati che non esiste alcuna correlazione tra il danno della cartilagine articolare e il dolore nei pazienti con OA [3,4] o modelli preclinici di OA [5], ha causato uno spostamento dell'attenzione per sviluppare farmaci per un efficace controllo del dolore . Questo articolo esaminerà le ultime scoperte nella ricerca sul dolore articolare ed evidenzierà alcuni degli obiettivi emergenti che potrebbero essere il futuro della gestione del dolore da artrite (riassunto in Fig. 1)
Le citochine
Le azioni di varie citochine negli studi di neurofisiologia articolare hanno avuto un ruolo piuttosto prominente di recente. L'interleuchina-6 (IL-6), per esempio, è una citochina che si lega tipicamente al recettore dell'IL-6 legato alla membrana (IL-6R). IL-6 può anche segnalare legandosi con un IL-6R solubile (SIL-6R) per produrre un complesso IL-6/sIL-6R. Questo complesso IL-6/sIL-6R si lega successivamente a una subunità glicoproteica transmembrana 130(gp130), consentendo così a IL-6 di segnalare in cellule che non esprimono costitutivamente IL-6R legato alla membrana [25,26]. IL-6 e SIL-6R sono attori chiave nell'infiammazione sistemica e nell'artrite, poiché è stata riscontrata una sovraregolazione di entrambi nel siero e nel liquido sinoviale dei pazienti affetti da AR [27,29]. Recentemente, Vazquez et al. hanno osservato che la co-somministrazione di IL-6/sIL-6R nelle ginocchia dei ratti ha causato dolore evocato da infiammazione, come rivelato da un aumento della risposta dei neuroni del corno dorsale spinale alla stimolazione meccanica del ginocchio e di altre parti. dell'arto posteriore [30]. L'ipereccitabilità dei neuroni spinali è stata osservata anche quando IL-6/sIL-6R è stato applicato localmente al midollo spinale. L'applicazione spinale di gp130 solubile (che assorbirebbe i complessi IL-6/sIL-6R, riducendo così la trans-segnalazione) ha inibito la sensibilizzazione centrale indotta da IL-6/sIL-6R. Tuttavia, l'applicazione acuta della sola gp130 solubile non ha ridotto le risposte neuronali all'infiammazione articolare già stabilita.
I canali del potenziale recettore transitorio (TRP) sono canali cationici non selettivi che agiscono come integratori di vari processi fisiologici e fisiopatologici. Oltre alla termosensazione, alla chemiosensazione e alla meccanosensazione, i canali TRP sono coinvolti nella modulazione del dolore e dell'infiammazione. Ad esempio, è stato dimostrato che i canali ionici TRP vanilloid-1 (TRPV1) contribuiscono al dolore infiammatorio articolare poiché l'iperalgesia termica non era evocabile nei topi monoartritici TRPV1 [31]. Allo stesso modo, i canali ionici TRP ankyrin-1 (TRPA1) sono coinvolti nell'ipersensibilità al meccano artritico poiché il blocco del recettore con antagonisti selettivi ha attenuato il dolore meccanico nell'infiammazione del modello adiuvante completo di Freunds [32,33]. Ulteriori prove che TRPV1 possa essere coinvolto nella neurotrasmissione del dolore da OA provengono da studi in cui l'espressione neuronale di TRPV1 è elevata nel modello di sodio monoiodoacetato di OA [34]. Inoltre, la somministrazione sistemica dell'antagonista TRPV1 A-889425 ha ridotto l'attività evocata e spontanea dell'intervallo dinamico spinale e dei neuroni specifici della nocicezione nel modello monoiodoacetato [35]. Questi dati suggeriscono che gli endovanilloidi potrebbero essere coinvolti nei processi di sensibilizzazione centrale associati al dolore da OA.
Attualmente sono noti almeno quattro polimorfismi nel gene che codifica per TRPV1, che portano ad un'alterazione nella struttura del canale ionico e ad una funzione compromessa. Un particolare polimorfismo (rs8065080) altera la sensibilità di TRPV1 alla capsaicina e gli individui portatori di questo polimorfismo sono meno sensibili all'iperalgesia termica [36]. Uno studio recente ha esaminato se i pazienti con OA con il polimorfismo rs8065080 hanno sperimentato un'alterata percezione del dolore basata su questa anomalia genetica. Il team di ricerca ha scoperto che i pazienti con OA asintomatica del ginocchio avevano maggiori probabilità di portare il gene rs8065080 rispetto ai pazienti con articolazioni doloranti [37]. Questa osservazione indica che i pazienti con OA con funzionamento normale; I canali TRPV1 hanno un aumentato rischio di dolore articolare e riaffermano il potenziale coinvolgimento di TRPV1 nella percezione del dolore OA.
Conclusione
Mentre l'ostacolo del trattamento del dolore da artrite rimane efficace, si stanno facendo grandi passi avanti nella nostra comprensione dei processi neurofisiologici responsabili della generazione del dolore articolare. Nuovi obiettivi vengono scoperti continuamente, mentre i meccanismi alla base dei percorsi noti vengono ulteriormente definiti e perfezionati. È improbabile che il targeting di uno specifico recettore o canale ionico sia la soluzione per normalizzare il dolore articolare, ma è piuttosto indicato un approccio polifarmaceutico in cui vari mediatori vengono utilizzati in combinazione durante fasi specifiche della malattia. Svelare i circuiti funzionali a ogni livello del percorso del dolore migliorerà anche la nostra conoscenza di come viene generato il dolore articolare. Ad esempio, identificare i mediatori periferici del dolore articolare ci consentirà di controllare la nocicezione all'interno dell'articolazione e probabilmente di evitare gli effetti collaterali centrali dei farmaci somministrati per via sistemica.
Dolore fetogenico
SINDROME DEL FACCIALE E DOLORE FACETOGENICO
Sindrome di Facet è un disturbo articolare correlato alle faccette articolari lombari e alle loro innervazioni e produce dolore facetogeno sia locale che radiante.
Un'eccessiva rotazione, estensione o flessione della colonna vertebrale (uso eccessivo ripetuto) può provocare alterazioni degenerative della cartilagine dell'articolazione. Inoltre, può comportare alterazioni degenerative di altre strutture, compreso il disco intervertebrale.
SINDROME DELLA FACCIA CERVICALE E DOLORE FACETOGENICO
Dolore al collo assiale (raramente che si irradia oltre le spalle), più comune unilateralmente.
Dolore con e / o limitazione di estensione e rotazione
Tenerezza dopo la palpazione
Irradiamento del dolore facetogenico localmente o nelle spalle o nella parte superiore della schiena, e raramente si irradia nella parte anteriore o in basso di un braccio o nelle dita come un ernia del disco potrebbe.
SINDROME DELLA FACCIA LOMBARE E DOLORE FACETOGENICO
Dolore o tenerezza nella parte bassa della schiena.
Tenerezza locale / rigidità a fianco della colonna vertebrale nella parte bassa della schiena.
Dolore, rigidità o difficoltà con alcuni movimenti (come stare in piedi o alzarsi da una sedia.
Dolore all'iperestensione
Il dolore riferito dalle articolazioni delle faccette lombari superiori può estendersi al fianco, all'anca e alla parte laterale superiore della coscia.
Il dolore riferito dalle faccette articolari lombari inferiori può penetrare in profondità nella coscia, lateralmente e/o posteriormente.
Le faccette articolari L4-L5 e L5-S1 possono riferirsi a dolore che si estende nella gamba laterale distale e, in rari casi, al piede
MEDICINA BASATA SULLE EVIDENZE
Medicina interventistica del dolore basata sull'evidenza secondo le diagnosi cliniche
12. Dolore originato dai lembi della faccetta lombare
Astratto
Sebbene l'esistenza di una sindrome delle faccette fosse stata a lungo messa in dubbio, ora è generalmente accettata come entità clinica. A seconda dei criteri diagnostici, le articolazioni zigapofisarie rappresentano tra il 5% e il 15% dei casi di lombalgia assiale cronica. Più comunemente, il dolore facetogeno deriva da stress ripetitivo e/o trauma cumulativo di basso livello, che porta a infiammazione e stiramento della capsula articolare. La lamentela più frequente è la lombalgia assiale con dolore riferito percepito al fianco, all'anca e alla coscia. Nessun risultato dell'esame obiettivo è patognomonico per la diagnosi. L'indicatore più forte per il dolore facetogeno lombare è la riduzione del dolore dopo blocchi anestetici dei rami mediali (rami mediali) dei rami dorsali che innervano le faccette articolari. Poiché possono verificarsi risultati falsi positivi e, possibilmente, falsi negativi, i risultati devono essere interpretati con attenzione. Nei pazienti con dolore articolare zigapofisario confermato dall'iniezione, gli interventi procedurali possono essere intrapresi nel contesto di un regime di trattamento multidisciplinare e multimodale che include farmacoterapia, terapia fisica ed esercizio fisico regolare e, se indicato, psicoterapia. Attualmente, il gold standard per il trattamento del dolore facetogeno è il trattamento con radiofrequenza (1 B+). Le prove a sostegno dei corticosteroidi intra-articolari sono limitate; quindi, questo dovrebbe essere riservato a coloro che non rispondono al trattamento con radiofrequenza (2 B1).
Il dolore facetogeno che emana dalle articolazioni delle faccette lombari è una causa comune di lombalgia nella popolazione adulta. Goldthwaite è stato il primo a descrivere la sindrome nel 1911 e Ghormley è generalmente accreditato per aver coniato il termine "sindrome delle faccette" nel 1933. Il dolore facetogeno è definito come il dolore che deriva da qualsiasi struttura che fa parte delle faccette articolari, inclusa la capsula fibrosa , membrana sinoviale, cartilagine ialina e osso.35
Più comunemente, è il risultato di stress ripetitivo e/o trauma cumulativo di basso livello. Ciò porta all'infiammazione, che può causare il riempimento e il rigonfiamento della faccetta articolare, con conseguente allungamento della capsula articolare e successiva generazione di dolore.27 I cambiamenti infiammatori intorno all'articolazione della faccetta possono anche irritare il nervo spinale attraverso il restringimento dei foraminali, con conseguente sciatica. Inoltre, Igarashi et al.28 hanno scoperto che le citochine infiammatorie rilasciate attraverso la capsula articolare ventrale in pazienti con degenerazione dell'articolazione zigapofisaria possono essere parzialmente responsabili dei sintomi neuropatici nei soggetti con stenosi spinale. I fattori predisponenti per il dolore articolare zigapofisario comprendono spondilolistesi / lisi, malattia degenerativa del disco e età avanzata. 5
IC PROVE AGGIUNTIVE
Il tasso di prevalenza di alterazioni patologiche nelle faccette articolari all'esame radiologico dipende dall'età media dei soggetti, dalla tecnica radiologica utilizzata e dalla definizione di anormalità. Le articolazioni delle faccette degenerative possono essere meglio visualizzate mediante esame di tomografia computerizzata (TC).49
DOLORE NEUROPATICO
Dolore iniziato o causato da una lesione o disfunzione primaria nel sistema nervoso somatosensoriale.
Il dolore neuropatico è solitamente cronico, difficile da trattare e spesso resistente alla gestione analgesica standard.
Astratto
Il dolore neuropatico è causato da una lesione o malattia del sistema somatosensoriale, comprese le fibre periferiche (fibre A?, A? e C) e i neuroni centrali, e colpisce il 7-10% della popolazione generale. Sono state descritte molteplici cause di dolore neuropatico. È probabile che la sua incidenza aumenti a causa dell'invecchiamento della popolazione globale, dell'aumento del diabete mellito e del miglioramento della sopravvivenza al cancro dopo la chemioterapia. Infatti, gli squilibri tra la segnalazione somatosensoriale eccitatoria e inibitoria, le alterazioni nei canali ionici e la variabilità nel modo in cui i messaggi del dolore sono modulati nel sistema nervoso centrale sono stati tutti implicati nel dolore neuropatico. Inoltre, l'onere del dolore neuropatico cronico sembra essere correlato alla complessità dei sintomi neuropatici, agli scarsi risultati e alle difficili decisioni terapeutiche. È importante sottolineare che la qualità della vita è compromessa nei pazienti con dolore neuropatico a causa dell'aumento delle prescrizioni di farmaci e delle visite agli operatori sanitari e della morbilità dovuta al dolore stesso e alla malattia scatenante. Nonostante le difficoltà, i progressi nella comprensione della fisiopatologia del dolore neuropatico stanno stimolando lo sviluppo di nuove procedure diagnostiche e interventi personalizzati, che sottolineano la necessità di un approccio multidisciplinare alla gestione del dolore neuropatico.
PATOGENESI DEL DOLORE NEUROPATICO
MECCANISMI PERIFERICI
Dopo una lesione del nervo periferico, i neuroni diventano più sensibili e sviluppano un'eccitabilità anormale e un'elevata sensibilità alla stimolazione.
Questo è noto come ... Sensibilizzazione periferica!
MECCANISMI CENTRALI
Come conseguenza dell'attività spontanea in corso nella periferia, i neuroni sviluppano un'attività di fondo aumentata, campi recettivi ampliati e risposte aumentate agli impulsi afferenti, compresi i normali stimoli tattili. Questo è noto come ... Sensibilizzazione centrale!
Il dolore neuropatico cronico è più frequente nelle donne (8% contro 5.7% negli uomini) e nei pazienti >50 anni (8.9% contro 5.6% in quelli <49 anni), e colpisce più comunemente la parte bassa della schiena e gli arti inferiori , collo e arti superiori24. Le radicolopatie dolorose lombari e cervicali sono probabilmente la causa più frequente di dolore neuropatico cronico. Coerentemente con questi dati, un'indagine su >12,000 pazienti con dolore cronico con tipi di dolore sia nocicettivo che neuropatico, indirizzata a specialisti del dolore in Germania, ha rivelato che il 40% di tutti i pazienti ha manifestato almeno alcune caratteristiche del dolore neuropatico (come sensazioni di bruciore, intorpidimento e formicolio); i pazienti con mal di schiena cronico e radicolopatia sono stati particolarmente colpiti25.
Il contributo della neurofisiologia clinica alla comprensione dei meccanismi di cefalea di tipo tensivo.
Astratto
Finora, gli studi neurofisiologici clinici sulla cefalea di tipo tensivo (TTH) sono stati condotti con due scopi principali: (1) stabilire se alcuni parametri neurofisiologici possono agire come marcatori di TTH e (2) studiare la fisiopatologia del TTH. Per quanto riguarda il primo punto, i risultati attuali sono deludenti poiché alcune anomalie riscontrate nei pazienti con TTH possono essere frequentemente osservate anche negli emicranici. D'altra parte, la neurofisiologia clinica ha svolto un ruolo importante nel dibattito sulla patogenesi del TTH. Gli studi sulla soppressione esterocettiva della contrazione del muscolo temporale hanno rilevato una disfunzione dell'eccitabilità del tronco cerebrale e del controllo soprasegmentale. Una conclusione simile è stata raggiunta utilizzando i riflessi trigeminocervicali, le cui anomalie nel TTH hanno suggerito una ridotta attività inibitoria degli interneuroni del tronco cerebrale, riflettendo meccanismi anormali di controllo del dolore endogeno. È interessante notare che l'anomalia dell'eccitabilità neurale nel TTH sembra essere un fenomeno generalizzato, non limitato ai distretti cranici. Meccanismi simil-DNIC difettosi sono stati infatti evidenziati anche nei distretti somatici da studi sul riflesso nocicettivo della flessione. Sfortunatamente, la maggior parte degli studi neurofisiologici sul TTH sono viziati da gravi difetti metodologici, che dovrebbero essere evitati in ricerche future per chiarire meglio i meccanismi del TTH.
Riferimenti:
Neurofisiologia del dolore da artrite. McDougall JJ1 Linton P.
Dolore che origina dalle articolazioni della faccetta lombare. van Kleef M1, Vanelderen P, Cohen SP, Lataster A, Van Zundert J, Mekhail N.
Il dolore neuropaticoLuana Colloca,1Taylor Ludman,1Didier Bouhassira,2Ralf Baron,3Anthony H. Dickenson,4David Yarnitsky,5Roy Freeman,6Andrea Truini,7Nadine Attal, Nanna B. Finnerup,9Christopher Eccleston,1011Eija Kalso,12David L. Bennett,13Robert H. Dworkin,14ed Srinivasa N. Raja15
Il contributo della neurofisiologia clinica alla comprensione dei meccanismi di cefalea di tipo tensivo. Rossi P1, Vollono C, Valeriani M, Sandrini G.
I medici definiscono il dolore cronico, come qualsiasi dolore che dura da 3 a 6 mesi o più. Il dolore influisce sulla salute mentale di un individuo e sulla vita quotidiana. Il dolore viene da una serie di messaggi che attraversano il sistema nervoso. La depressione sembra seguire il dolore. Provoca gravi sintomi che influenzano il modo in cui un individuo sente, pensa e come gestisce le attività quotidiane, cioè dormire, mangiare e lavorare. Chiropratico, il Dr. Alex Jimenez approfondisce potenziali biomarcatori che possono aiutare a trovare e curare le cause alla radice del dolore e del dolore cronico.
Il primo passo nella gestione del dolore di successo è una valutazione biopsicosociale completa.
L'estensione della patologia organica potrebbe non riflettersi con precisione nell'esperienza del dolore.
La valutazione iniziale può essere utilizzata per identificare le aree che richiedono una valutazione più approfondita.
Sono disponibili molti strumenti autovalutati validati per valutare l'impatto del dolore cronico.
Valutazione dei pazienti con dolore cronico
Il dolore cronico è un problema di salute pubblica che colpisce il 20-30% della popolazione dei paesi occidentali. Sebbene ci siano stati molti progressi scientifici nella comprensione della neurofisiologia del dolore, la valutazione e la diagnosi precise del problema di dolore cronico di un paziente non sono né chiare né ben definite. Il modo in cui il dolore cronico viene concettualizzato influenza il modo in cui il dolore viene valutato e i fattori considerati durante la diagnosi del dolore cronico. Non esiste una relazione uno-a-uno tra la quantità o il tipo di patologia organica e l'intensità del dolore, ma invece, l'esperienza del dolore cronico è modellata da una miriade di aspetti biomedici, psicosociali (ad es. convinzioni, aspettative e umore dei pazienti) e fattori comportamentali (ad es. contesto, risposte di altri significativi). Valutare ciascuno di questi tre domini attraverso una valutazione completa della persona con dolore cronico è essenziale per le decisioni terapeutiche e per facilitare i risultati ottimali. Questa valutazione dovrebbe includere una storia completa del paziente e una valutazione medica e un breve colloquio di screening in cui il comportamento del paziente può essere osservato. Un'ulteriore valutazione per affrontare le questioni identificate durante la valutazione iniziale guiderà le decisioni su quali eventuali ulteriori valutazioni potrebbero essere appropriate. Sono disponibili strumenti standardizzati autovalutati per valutare l'intensità del dolore, le capacità funzionali, le convinzioni e le aspettative e il disagio emotivo del paziente, che possono essere somministrati dal medico o un referente per una valutazione approfondita che può essere di aiuto nella pianificazione del trattamento.
Il dolore è un sintomo estremamente diffuso. Si stima che il solo dolore cronico influenzi il 30% della popolazione adulta degli Stati Uniti, in aumento di 100 milioni di adulti.1
Nonostante il costo elevato di trattare le persone con dolore cronico, il sollievo per molti rimane elusivo e la completa eliminazione del dolore è rara. Sebbene ci siano stati progressi sostanziali nella conoscenza della neurofisiologia del dolore, insieme allo sviluppo di potenti farmaci analgesici e altri interventi medici e chirurgici innovativi, in media la quantità di riduzione del dolore con le procedure disponibili è del 30-40% e ciò si verifica in meno della metà dei pazienti trattati.
Il modo in cui pensiamo al dolore influenza il modo in cui valutiamo il dolore. La valutazione inizia con la storia e l'esame obiettivo, seguita da test di laboratorio e procedure di diagnostica per immagini nel tentativo di identificare e / o confermare la presenza di qualsiasi patologia sottostante che causa il / i sintomo / i generatore di dolore.
In assenza di patologia organica identificabile, l'operatore sanitario può presumere che il rapporto dei sintomi deriva da fattori psicologici e può richiedere una valutazione psicologica per rilevare i fattori emotivi alla base del rapporto del paziente. C'è dualità in cui il rapporto dei sintomi è attribuito a entrambi somatico or meccanismi psicogeni.
Ad esempio, le basi organiche per alcuni dei più comuni e ricorrenti acuti (ad es. Mal di testa) 3 e cronici [ad es. mal di schiena, fibromialgia (FM)] i problemi di dolore sono in gran parte sconosciuti, 4,5 mentre d'altra parte, gli individui asintomatici possono avere anomalie strutturali come ernia del disco che spiegherebbe il dolore se fosse presente.6,7 Mancano spiegazioni adeguate per i pazienti senza patologia organica identificata che segnalano dolore severo e individui indolori con patologia oggettiva e significativa.
Il dolore cronico colpisce non solo il singolo paziente, ma anche i suoi altri significativi (partner, parenti, datori di lavoro, colleghi di lavoro e amici), rendendo essenziale un trattamento appropriato. Un trattamento soddisfacente può venire solo dalla valutazione completa dell'eziologia biologica del dolore in concomitanza con la specifica presentazione psicosociale e comportamentale del paziente, incluso il loro stato emotivo (ad esempio ansia, depressione e rabbia), percezione e comprensione dei sintomi e reazioni a quelli sintomi di altri significativi.8,9 Una premessa chiave è che più fattori influenzano i sintomi e i limiti funzionali degli individui con dolore cronico. Pertanto, è necessaria una valutazione completa che affronta domini biomedici, psicosociali e comportamentali, poiché ciascuno contribuisce al dolore cronico e alla disabilità correlata. 10,11
Valutazione completa di un individuo con dolore cronico
Turk e Meichenbaum12 hanno suggerito che tre domande centrali dovrebbero guidare la valutazione delle persone che riferiscono di dolore:
Qual è l'entità della malattia o della lesione del paziente (menomazione fisica)?
Qual è la grandezza della malattia? Cioè, fino a che punto il paziente soffre, è disabile e non è in grado di godersi le solite attività?
Il comportamento dell'individuo sembra appropriato alla malattia o alla lesione, oppure esistono prove dell'amplificazione dei sintomi per una serie di motivi psicologici o sociali (ad esempio benefici quali attenzione positiva, farmaci che alterano l'umore, compensazione finanziaria)?
Per rispondere a queste domande, le informazioni dovrebbero essere raccolte dal paziente mediante anamnesi ed esame obiettivo, in combinazione con un colloquio clinico e attraverso strumenti di valutazione standardizzati. Gli operatori sanitari devono cercare qualsiasi causa di dolore attraverso l'esame obiettivo e i test diagnostici mentre valutano contemporaneamente l'umore, le paure, le aspettative, gli sforzi di coping del paziente, le risorse, le risposte di altre persone significative e l'impatto del dolore sui pazienti. vite.11 In breve, l'operatore sanitario deve valutare la `` persona intera '' e non solo il dolore.
Gli obiettivi generali della storia e della valutazione medica sono:
(i) determinare la necessità di ulteriori test diagnostici
(ii) determinare se i dati medici possono spiegare i sintomi del paziente, la gravità dei sintomi e i limiti funzionali
(iii) fare una diagnosi medica
(iv) valutare la disponibilità di un trattamento appropriato
(v) stabilire gli obiettivi del trattamento
(vi) determinare il corso appropriato per la gestione dei sintomi se non è possibile una cura completa.
Un numero significativo di pazienti che riferiscono di dolore cronico non dimostrano alcuna patologia fisica utilizzando radiografie semplici, scansioni tomografiche assiali calcolate o elettromiografia (una vasta letteratura è disponibile sulla valutazione fisica, sulle procedure di valutazione radiologica e di laboratorio per determinare le basi fisiche del dolore), 17 rende difficile o impossibile una diagnosi patologica precisa.
Nonostante queste limitazioni, la storia e l'esame fisico del paziente rimangono alla base della diagnosi medica, possono fornire una salvaguardia contro i risultati sovrainterpretativi dall'imaging diagnostico che sono ampiamente confermativi e possono essere utilizzati per guidare la direzione di ulteriori sforzi di valutazione.
Inoltre, i pazienti con problemi di dolore cronico spesso consumano una varietà di farmaci.18 È importante discutere gli attuali farmaci di un paziente durante l'intervista, poiché molti farmaci antidolorifici sono associati ad effetti collaterali che possono causare o simulare un disagio emotivo.19 Gli operatori sanitari dovrebbero non solo avere familiarità con i farmaci usati per il dolore cronico, ma anche con gli effetti collaterali di questi farmaci che si traducono in affaticamento, difficoltà di sonno e cambiamenti dell'umore per evitare diagnosi errate della depressione.
Si ritiene che l'uso dei diari quotidiani sia più accurato in quanto sono basati sul tempo reale piuttosto che sul richiamo. Ai pazienti può essere chiesto di mantenere regolari i diari dell'intensità del dolore con valutazioni registrate più volte al giorno (ad esempio, pasti e prima di coricarsi) per diversi giorni o settimane e le valutazioni del dolore multiple possono essere mediate nel tempo.
Un problema notato con l'uso di diari di carta e matita è che i pazienti potrebbero non seguire le istruzioni per fornire valutazioni a intervalli specificati. Piuttosto, i pazienti possono completare i diari in anticipo ("riempire in avanti") o poco prima di vedere un medico ("riempire all'indietro"), 24 minando la presunta validità dei diari. I diari elettronici hanno ottenuto l'accettazione in alcuni studi di ricerca per evitare questi problemi.
La ricerca ha dimostrato l'importanza di valutare la qualità della vita complessiva correlata alla salute (HRQOL) in pazienti con dolore cronico oltre alla funzione.31,32 Esistono numerose misure HRQOL ben consolidate e supportate psicometricamente [Indagine sulla salute a breve termine dello studio sui risultati medici (SF) -36)], 33 misure generali del funzionamento fisico [ad es. Indice di disabilità del dolore (PDI)], 34 e misure specifiche per la malattia [ad es. Indice di osteoartrosi Western Ontario MacMaster (WOMAC); 35 Roland-Morris Back Pain Disability Questionnaire (RDQ)] 36 per valutare la funzione e la qualità della vita.
Le misure specifiche per la malattia sono progettate per valutare l'impatto di una condizione specifica (ad esempio dolore e rigidità nelle persone con osteoartrite), mentre le misure generiche consentono di confrontare il funzionamento fisico associato a un dato disturbo e il suo trattamento con quello di varie altre condizioni. Gli effetti specifici di un disturbo possono non essere rilevati quando si utilizza una misura generica; pertanto, le misure specifiche per la malattia possono essere più probabili per rivelare un miglioramento clinicamente importante o un deterioramento delle funzioni specifiche come risultato del trattamento. Misure generali di funzionamento possono essere utili per confrontare i pazienti con una varietà di condizioni dolorose. L'uso combinato di misure specifiche per malattia e generiche facilita il raggiungimento di entrambi gli obiettivi.
La presenza di disagio emotivo nelle persone con dolore cronico rappresenta una sfida nel valutare sintomi come affaticamento, riduzione del livello di attività, diminuzione della libido, alterazione dell'appetito, disturbi del sonno, aumento o perdita di peso e deficit di memoria e concentrazione, poiché questi sintomi possono essere risultato di dolore, sofferenza emotiva o trattamento prescritto per controllare il dolore.
Gli strumenti sono stati sviluppati specificamente per i pazienti affetti da dolore per valutare il disagio psicologico, l'impatto del dolore sulla vita dei pazienti, la sensazione di controllo, i comportamenti di coping e gli atteggiamenti nei confronti della malattia, del dolore e degli operatori sanitari.17
Ad esempio, il Beck Depression Inventory (BDI) 39 e il Profile of Mood States (POMS) 40 sono psicometrici per valutare i sintomi di umore depresso, disagio emotivo e disturbi dell'umore e sono stati raccomandati per essere utilizzati in tutti gli studi clinici di dolore cronico, 41 tuttavia, i punteggi devono essere interpretati con cautela e potrebbe essere necessario modificare i criteri per i livelli di stress emotivo per evitare falsi positivi.42
Biomarker del laboratorio per il dolore
I biomarcatori sono caratteristiche biologiche che possono essere utilizzate per indicare la salute o la malattia. Questo articolo esamina gli studi sui biomarcatori della lombalgia (LBP) in soggetti umani. La LBP è la principale causa di disabilità, causata da vari disturbi correlati alla colonna vertebrale, tra cui la degenerazione del disco intervertebrale, l'ernia del disco, la stenosi spinale e l'artrite della faccetta. Il focus di questi studi è quello dei mediatori dell'infiammazione, perché l'infiammazione contribuisce alla patogenesi della degenerazione del disco e dei meccanismi associati al dolore. Sempre più spesso, gli studi suggeriscono che la presenza di mediatori dell'infiammazione può essere misurata sistematicamente nel sangue. Questi biomarcatori possono servire come nuovi strumenti per dirigere la cura del paziente. Attualmente, la risposta del paziente al trattamento è imprevedibile con un tasso significativo di recidiva e, mentre i trattamenti chirurgici possono fornire la correzione anatomica e il sollievo dal dolore, sono invasivi e costosi. La revisione comprende studi condotti su popolazioni con diagnosi specifiche e origini indefinite di LBP. Poiché la storia naturale della LBP è progressiva, la natura temporale degli studi è categorizzata dalla durata della sintomatologia / malattia. Vengono inoltre esaminati studi correlati sui cambiamenti nei biomarcatori con trattamento. In definitiva, i biomarcatori diagnostici della LBP e della degenerazione spinale hanno il potenziale per condurre un'era di medicina della colonna vertebrale individualizzata per terapie personalizzate nel trattamento della LBP.
Biomarcatori per dolore neuropatico cronico e potenziale applicazione nella stimolazione del midollo spinale
Questa recensione si è concentrata sulla comprensione di quali sostanze all'interno del corpo umano aumentano e diminuiscono con l'aumentare del dolore neuropatico. Abbiamo esaminato vari studi e abbiamo visto correlazioni tra dolore neuropatico e componenti del sistema immunitario (questo sistema difende il corpo da malattie e infezioni). I nostri risultati saranno particolarmente utili per capire come ridurre o eliminare il disagio, il dolore neuropatico cronico porta con sé. La procedura di stimolazione del midollo spinale (SCS) è uno dei pochi trattamenti correttivi abbastanza efficaci per il dolore. Uno studio di follow-up applicherà i risultati di questa revisione a SCS, al fine di comprendere il meccanismo e ottimizzare ulteriormente l'efficacia.
È stato scoperto che le citochine proinfiammatorie come IL-1, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 e TNF-? Svolgono un ruolo significativo nell'amplificazione degli stati di dolore cronico.
Dopo la revisione di vari studi relativi ai biomarcatori del dolore, abbiamo scoperto che i livelli sierici di citochine e chemochine pro-infiammatorie, come IL-1 ?, IL-6, IL-2, IL-33, CCL3, CXCL1, CCR5 e TNF - ?, erano significativamente up-regolati durante l'esperienza del dolore cronico. D'altra parte, è stato scoperto che le citochine antinfiammatorie come IL-10 e IL-4 mostrano una significativa down-regolazione durante lo stato di dolore cronico.
Biomarcatori per la depressione
Una pletora di ricerche ha implicato centinaia di putativi biomarcatori per la depressione, ma non ha ancora chiarito completamente i loro ruoli nella malattia depressiva o stabilito ciò che è anormale in cui i pazienti e in che modo le informazioni biologiche possono essere utilizzate per migliorare diagnosi, trattamento e prognosi. Questa mancanza di progresso è parzialmente dovuta alla natura e all'eterogeneità della depressione, in combinazione con l'eterogeneità metodologica all'interno della letteratura di ricerca e la vasta gamma di biomarcatori con potenziale, la cui espressione varia spesso in base a molti fattori. Esaminiamo la letteratura disponibile, che indica che i marcatori coinvolti nei processi infiammatori, neurotrofici e metabolici, così come i componenti del sistema neurotrasmettitore e neuroendocrino, rappresentano candidati molto promettenti. Questi possono essere misurati attraverso valutazioni genetiche ed epigenetiche, trascrittomiche e proteomiche, metabolomiche e di neuroimaging. L'uso di nuovi approcci e programmi di ricerca sistematici è ora necessario per determinare se e quali biomarcatori possono essere utilizzati per prevedere la risposta al trattamento, stratificare i pazienti in trattamenti specifici e sviluppare obiettivi per nuovi interventi. Concludiamo che ci sono molte promesse per ridurre il peso della depressione attraverso l'ulteriore sviluppo e l'espansione di questi percorsi di ricerca.
Riferimenti:
Valutazione dei pazienti con dolore cronico EJ Dansiet e DC Turk * t
Biomarcatori infiammatori di lombalgia e degenerazione del disco: una review.
Khan AN1, Jacobsen HE2, Khan J1, Filippi CG3, Levine M3, Lehman RA Jr2,4, Riew KD2,4, Lenke LG2,4, Chahine NO2,5.
Biomarcatori per il dolore neuropatico cronico e la loro potenziale applicazione nella stimolazione del midollo spinale: una revisione
Chibueze D. Nwagwu, 1 Christina Sarris, MD, 3 Yuan-Xiang Tao, Ph.D., MD, 2 e Antonios Mammis, MD1,2
Biomarcatori per la depressione: approfondimenti recenti, sfide attuali e prospettive future. Strawbridge R1, Young AH1,2, Cleare AJ1,2.
Il dolore è la naturale risposta del corpo umano a lesioni o malattie, ed è spesso un avvertimento che qualcosa non va. Una volta che il problema è guarito, generalmente smettiamo di sperimentare questi sintomi dolorosi, tuttavia, cosa succede quando il dolore persiste molto tempo dopo che la causa è sparita? Dolore cronico è definito medico come dolore persistente che dura da 3 a 6 mesi o più. Il dolore cronico è certamente una condizione difficile da vivere con, che interessano tutto, dai livelli di attività del soggetto e la loro capacità di lavorare così come le loro relazioni personali e le condizioni psicologiche. Ma sei consapevole che il dolore cronico può anche influenzare la struttura e la funzione del tuo cervello? Si scopre che questi cambiamenti cerebrali possono portare a problemi cognitivi e psicologici.
Il dolore cronico non influenza solo una singolare regione della mente, in quanto può portare a cambiamenti in numerose aree essenziali del cervello, molte delle quali sono coinvolte in molti processi e funzioni fondamentali. Vari studi di ricerca nel corso degli anni hanno trovato alterazioni dell'ippocampo, insieme alla riduzione della materia grigia dalla corteccia prefrontale dorsolaterale, dall'amigdala, dal tronco cerebrale e dalla corteccia insulare destra, solo per citarne alcuni, associati al dolore cronico. Una ripartizione di alcune delle strutture di queste regioni e delle loro relative funzioni potrebbe aiutare a mettere questi cambiamenti cerebrali nel contesto, per un sacco di individui con dolore cronico. Lo scopo del seguente articolo è quello di dimostrare e discutere i cambiamenti cerebrali strutturali e funzionali associati al dolore cronico, in particolare nel caso in cui quelli riflettono probabilmente né danno né atrofia.
I cambiamenti strutturali del cervello nel dolore cronico riflettono probabilmente né danno né atrofia
Astratto
Il dolore cronico sembra essere associato alla riduzione della materia grigia cerebrale in aree ascrivibili alla trasmissione del dolore. I processi morfologici alla base di questi cambiamenti strutturali, probabilmente in seguito alla riorganizzazione funzionale e alla plasticità centrale del cervello, rimangono poco chiari. Il dolore nell'osteoartrosi dell'anca è una delle poche sindromi dolorose croniche che sono principalmente curabili. Abbiamo studiato 20 pazienti con dolore cronico dovuto a coxartrosi unilaterale (età media 63.25-9.46 (DS) anni, 10 femmine) prima della chirurgia endoprotetica dell'articolazione dell'anca (stato doloroso) e monitorato i cambiamenti strutturali del cervello fino a 1 anno dopo l'intervento: 6-8 settimane , 12-18 settimane e 10-14 mesi quando è completamente indolore. I pazienti con dolore cronico dovuto a coxartrosi unilaterale avevano una sostanza grigia significativamente inferiore rispetto ai controlli nella corteccia cingolata anteriore (ACC), nella corteccia insulare e nell'opercolo, nella corteccia prefrontale dorsolaterale (DLPFC) e nella corteccia orbitofrontale. Queste regioni funzionano come strutture multi-integrative durante l'esperienza e l'anticipazione del dolore. Quando i pazienti erano indolori dopo il recupero dalla chirurgia endoprotetica, è stato riscontrato un aumento della sostanza grigia in quasi le stesse aree. Abbiamo anche riscontrato un progressivo aumento della materia grigia cerebrale nella corteccia premotoria e nell'area motoria supplementare (SMA). Concludiamo che le anomalie della materia grigia nel dolore cronico non sono la causa, ma sono secondarie alla malattia e sono almeno in parte dovute a cambiamenti nella funzione motoria e nell'integrazione corporea.
Introduzione
Le prove di riorganizzazione funzionale e strutturale nei pazienti con dolore cronico supportano l'idea che il dolore cronico non dovrebbe essere concettualizzato solo come uno stato funzionale alterato, ma anche come conseguenza della plasticità cerebrale funzionale e strutturale [1], [2], [3], [4], [5], [6]. Negli ultimi sei anni sono stati pubblicati più di 20 studi che dimostrano i cambiamenti strutturali del cervello in 14 sindromi da dolore cronico. Una caratteristica sorprendente di tutti questi studi è il fatto che i cambiamenti della materia grigia non sono stati distribuiti in modo casuale, ma si verificano in aree cerebrali definite e funzionalmente altamente specifiche, vale a dire, il coinvolgimento nell'elaborazione nocicettiva sopraspinale. I risultati più importanti erano diversi per ciascuna sindrome dolorosa, ma si sovrapponevano nella corteccia cingolata, nella corteccia orbitofrontale, nell'isola e nel ponte dorsale [4]. Ulteriori strutture comprendono il talamo, la corteccia prefrontale dorsolaterale, i gangli della base e l'area dell'ippocampo. Questi risultati sono spesso discussi come atrofia cellulare, rafforzando l'idea di danno o perdita di materia grigia cerebrale [7], [8], [9]. In effetti, i ricercatori hanno trovato una correlazione tra la diminuzione della materia grigia cerebrale e la durata del dolore [6], [10]. Ma la durata del dolore è anche collegata all'età del paziente e il declino globale della materia grigia dipendente dall'età, ma anche regionale specifico, è ben documentato [11]. D'altra parte, questi cambiamenti strutturali potrebbero anche essere una diminuzione delle dimensioni delle cellule, fluidi extracellulari, sinaptogenesi, angiogenesi o anche a causa di cambiamenti del volume sanguigno [4], [12], [13]. Qualunque sia la fonte, per la nostra interpretazione di tali risultati è importante vedere questi risultati morfometrici alla luce di una vasta gamma di studi morfometrici sulla plasticità dipendente dall'esercizio, dato che i cambiamenti cerebrali strutturali specifici a livello regionale sono stati ripetutamente mostrati a seguito di esercizio cognitivo e fisico [ 14].
Non si capisce perché solo una percentuale relativamente piccola di esseri umani sviluppa una sindrome del dolore cronico, considerando che il dolore è un'esperienza universale. Sorge la domanda se in alcuni esseri umani una differenza strutturale nei sistemi di trasmissione del dolore centrale possa agire come una diatesi per il dolore cronico. I cambiamenti della materia grigia nel dolore fantasma dovuto all'amputazione [15] e alla lesione del midollo spinale [3] indicano che i cambiamenti morfologici del cervello sono, almeno in parte, una conseguenza del dolore cronico. Tuttavia, il dolore nell'osteoartrosi dell'anca (OA) è una delle poche sindrome da dolore cronico che è principalmente curabile, poiché l'88% di questi pazienti è regolarmente libero dal dolore dopo un intervento di sostituzione totale dell'anca (THR) [16]. In uno studio pilota abbiamo analizzato dieci pazienti con OA dell'anca prima e subito dopo l'intervento chirurgico. Abbiamo riscontrato diminuzioni di materia grigia nella corteccia cingolata anteriore (ACC) e nell'insula durante il dolore cronico prima dell'intervento THR e abbiamo riscontrato aumenti di materia grigia nelle corrispondenti aree cerebrali nella condizione senza dolore dopo l'intervento chirurgico [17]. Concentrandoci su questo risultato, ora abbiamo ampliato i nostri studi indagando su più pazienti (n? =? 20) dopo THR di successo e monitorato i cambiamenti strutturali del cervello in quattro intervalli di tempo, fino a un anno dopo l'intervento chirurgico. Per controllare i cambiamenti della materia grigia dovuti al miglioramento motorio o alla depressione, abbiamo anche somministrato questionari mirati al miglioramento della funzione motoria e della salute mentale.
Materiali e Metodi
Volontari
I pazienti qui riportati sono un sottogruppo di 20 pazienti su 32 pazienti pubblicati di recente che sono stati confrontati con un gruppo di controllo sano di pari età e sesso [17] ma hanno partecipato a un'ulteriore indagine di follow-up di un anno. Dopo l'intervento chirurgico 12 pazienti hanno abbandonato la scuola a causa di un secondo intervento di chirurgia endoprotetica (n? =? 2), malattia grave (n? =? 2) e revoca del consenso (n? =? 8). Ciò ha lasciato un gruppo di venti pazienti con OA primaria unilaterale dell'anca (età media 63.25-9.46 (DS) anni, 10 femmine) che sono stati esaminati quattro volte: prima dell'intervento chirurgico (stato doloroso) e ancora 6 8 e 12 18 settimane e 10 14 mesi dopo la chirurgia endoprotetica, quando completamente indolore. Tutti i pazienti con OA primaria dell'anca avevano un'anamnesi dolorosa superiore a 12 mesi, compresa tra 1 e 33 anni (media 7.35 anni) e un punteggio medio del dolore di 65.5 (compreso tra 40 e 90) su una scala analogica visiva (VAS) compresa tra Da 0 (nessun dolore) a 100 (peggior dolore immaginabile). Abbiamo valutato qualsiasi evento di dolore minore, inclusi mal di denti, orecchie e cefalea fino a 4 settimane prima dello studio. Abbiamo anche selezionato casualmente i dati da 20 controlli sani abbinati per sesso ed età (età media 60,95 8,52 (SD) anni, 10 femmine) dei 32 dello studio pilota sopra menzionato [17]. Nessuno dei 20 pazienti o dei 20 volontari sani di pari età e sesso aveva una storia medica interna o neurologica. Lo studio ha ricevuto l'approvazione etica dal comitato etico locale e il consenso informato scritto è stato ottenuto da tutti i partecipanti allo studio prima dell'esame.
Dati comportamentali
Abbiamo raccolto dati su depressione, somatizzazione, ansia, dolore e salute fisica e mentale in tutti i pazienti e in tutti e quattro i punti temporali utilizzando i seguenti questionari standardizzati: Beck Depression Inventory (BDI) [18], Brief Symptom Inventory (BSI) [19], Schmerzempfindungs-Skala (SES? =? Scala di spiacevolezza del dolore) [20] e Health Survey 36-Item Short Form (SF-36) [21] e Nottingham Health Profile (NHP). Abbiamo condotto misure ripetute ANOVA e test t a due code accoppiati per analizzare i dati comportamentali longitudinali utilizzando SPSS 13.0 per Windows (SPSS Inc., Chicago, IL) e abbiamo utilizzato la correzione Greenhouse Geisser se il presupposto per la sfericità è stato violato. Il livello di significatività è stato fissato a p <0.05.
VBM - Acquisizione dati
Acquisizione dell'immagine. La scansione RM ad alta risoluzione è stata eseguita su un sistema MRI 3T (Siemens Trio) con una bobina per la testa a 12 canali standard. Per ciascuno dei quattro punti temporali, scansione I (tra 1 giorno e 3 mesi prima della chirurgia endoprotetica), scansione II (da 6 a 8 settimane dopo l'intervento), scansione III (da 12 a 18 settimane dopo l'intervento) e scansione IV (10-14 mesi dopo l'intervento), è stata acquisita una risonanza magnetica strutturale T1 pesata per ciascun paziente utilizzando una sequenza 3D-FLASH (TR 15 ms, TE 4.9 ms, angolo di vibrazione 25 , fette di 1 mm, FOV 256, dimensione voxel 256 1 1 mm).
Elaborazione delle immagini e analisi statistica
La pre-elaborazione e l'analisi dei dati sono state eseguite con SPM2 (Wellcome Department of Cognitive Neurology, Londra, UK) in esecuzione sotto Matlab (Mathworks, Sherborn, MA, USA) e contenente una morfometria basata su voxel (VBM) -toolbox per dati longitudinali, che si basa su immagini MR 3D strutturali ad alta risoluzione e consente di applicare statistiche voxel per rilevare le differenze regionali nella densità o nei volumi della materia grigia [22], [23]. In sintesi, la pre-elaborazione ha coinvolto la normalizzazione spaziale, la segmentazione della materia grigia e il livellamento spaziale di 10 mm con un kernel gaussiano. Per le fasi di pre-elaborazione, abbiamo utilizzato un protocollo ottimizzato [22], [23] e un modello di materia grigia specifico per scanner e studio [17]. Abbiamo usato SPM2 piuttosto che SPM5 o SPM8 per rendere questa analisi paragonabile al nostro studio pilota [17]. in quanto consente un'ottima normalizzazione e segmentazione dei dati longitudinali. Tuttavia, poiché un aggiornamento più recente di VBM (VBM8) è diventato disponibile di recente (dbm.neuro.uni-jena.de/vbm/), abbiamo utilizzato anche VBM8.
Analisi cross-sectional
Abbiamo utilizzato un test t a due campioni per rilevare le differenze regionali nella materia grigia del cervello tra i gruppi (pazienti al punto temporale I (dolore cronico) e controlli sani). Abbiamo applicato una soglia di p <0.001 (non corretta) in tutto il cervello a causa della nostra forte ipotesi di priorità, che si basa su 9 studi indipendenti e coorti che mostrano una diminuzione della materia grigia nei pazienti con dolore cronico [7], [8], [ 9], [15], [24], [25], [26], [27], [28], che gli aumenti di materia grigia appariranno nelle stesse regioni (rilevanti per l'elaborazione del dolore) del nostro studio pilota (17 ). I gruppi sono stati abbinati per età e sesso senza differenze significative tra i gruppi. Per indagare se le differenze tra i gruppi sono cambiate dopo un anno, abbiamo anche confrontato i pazienti alla scansione del punto temporale IV (senza dolore, un anno di follow-up) con il nostro gruppo di controllo sano.
Analisi longitudinale
Per rilevare le differenze tra i punti temporali (Scan I IV) abbiamo confrontato le scansioni prima dell'intervento chirurgico (stato del dolore) e di nuovo 6 8 e 12 18 settimane e 10 14 mesi dopo la chirurgia endoprotetica (indolore) come misura ripetuta ANOVA. Poiché qualsiasi cambiamento cerebrale dovuto al dolore cronico può richiedere un po 'di tempo per regredire dopo l'operazione e la cessazione del dolore ea causa del dolore post-operatorio riportato dai pazienti, abbiamo confrontato la scansione longitudinale I e II con la scansione III e IV. Per rilevare i cambiamenti che non sono strettamente collegati al dolore, abbiamo anche cercato cambiamenti progressivi su tutti gli intervalli di tempo. Abbiamo capovolto il cervello dei pazienti con OA dell'anca sinistra (n? =? 7) al fine di normalizzare il lato del dolore per entrambi, il confronto di gruppo e l'analisi longitudinale, ma principalmente abbiamo analizzato i dati non ribaltati. Abbiamo utilizzato il punteggio BDI come covariata nel modello.
Risultati
Dati comportamentali
Tutti i pazienti hanno riportato dolore cronico all'anca prima dell'intervento chirurgico ed erano indolori (per quanto riguarda questo dolore cronico) immediatamente dopo l'intervento, ma hanno riportato un dolore post-operatorio piuttosto acuto alla scansione II che era diverso dal dolore dovuto all'osteoartrosi. Il punteggio di salute mentale dell'SF-36 (F (1.925 / 17.322)? =? 0.352, p? =? 0.7) e il punteggio globale BSI GSI (F (1.706 / 27.302)? =? 3.189, p? =? 0.064 ) non hanno mostrato cambiamenti nel corso del tempo e nessuna comorbilità mentale. Nessuno dei controlli ha riportato alcun dolore acuto o cronico e nessuno ha mostrato sintomi di depressione o disabilità fisica / mentale.
Prima dell'intervento chirurgico, alcuni pazienti mostravano sintomi depressivi da lievi a moderati nei punteggi BDI che diminuivano significativamente alla scansione III (t (17)? =? 2.317, p? =? 0.033) e IV (t (16)? =? 2.132, p? =? 0.049). Inoltre, i punteggi SES (spiacevolezza del dolore) di tutti i pazienti sono migliorati significativamente dalla scansione I (prima dell'intervento) alla scansione II (t (16)? =? 4.676, p <0.001), scansione III (t (14)? =? 4.760, p <0.001) e scansione IV (t (14)? =? 4.981, p <0.001, 1 anno dopo l'intervento chirurgico) poiché la sgradevolezza del dolore diminuiva con l'intensità del dolore. La valutazione del dolore nelle scansioni 1 e 2 è stata positiva, la stessa valutazione nei giorni 3 e 4 negativa. Il tutor descrive solo la qualità del dolore percepito. È stato quindi positivo il giorno 1 e 2 (media 19.6 il giorno 1 e 13.5 il giorno 2) e negativo (na) il giorno 3 e 4. Tuttavia, alcuni pazienti non hanno compreso questa procedura e hanno utilizzato il SES come qualità globale della misura della vita. Questo è il motivo per cui a tutti i pazienti è stato chiesto lo stesso giorno individualmente e dalla stessa persona in merito alla comparsa del dolore.
Nel sondaggio sulla salute in forma abbreviata (SF-36), che consiste nelle misure di riepilogo di un punteggio di salute fisica e un punteggio di salute mentale [29], i pazienti sono migliorati significativamente nel punteggio di salute fisica dalla scansione I alla scansione II (t ( 17)? = ?? 4.266, p? =? 0.001), scansione III (t (16)? = ?? 8.584, p <0.001) e IV (t (12)? = ?? 7.148, p <0.001), ma non nel punteggio della salute mentale. I risultati del NHP erano simili, nella sottoscala `` dolore '' (polarità invertita) abbiamo osservato un cambiamento significativo dalla scansione I alla scansione II (t (14)? = ?? 5.674, p <0.001, scansione III (t (12 )? = ?? 7.040, p <0.001 e scansione IV (t (10)? = ?? 3.258, p? =? 0.009). Abbiamo anche riscontrato un aumento significativo della sottoscala `` mobilità fisica '' dalla scansione I alla scansione III (t (12)? = ?? 3.974, p? =? 0.002) e scansione IV (t (10)? = ?? 2.511, p? =? 0.031). Non c'è stato alcun cambiamento significativo tra la scansione I e la scansione II ( sei settimane dopo l'intervento).
Dati strutturali
Analisi trasversale. Abbiamo incluso l'età come covariata nel modello lineare generale e non abbiamo trovato fattori di confusione relativi all'età. Rispetto ai controlli di pari età e sesso, i pazienti con OA primaria dell'anca (n? =? 20) hanno mostrato preoperatoriamente (Scan I) una riduzione della materia grigia nella corteccia cingolata anteriore (ACC), nella corteccia insulare, nell'opercolo, nella corteccia prefrontale dorsolaterale ( DLPFC), polo temporale destro e cervelletto (Tabella 1 e Figura 1). Ad eccezione del putamen destro (x? =? 31, y? = ?? 14, z? = ?? 1; p <0.001, t? =? 3.32) non è stato riscontrato alcun aumento significativo della densità della sostanza grigia nei pazienti con OA rispetto a controlli sani. Confrontando i pazienti alla scansione del punto temporale IV con i controlli abbinati, sono stati trovati gli stessi risultati dell'analisi della sezione trasversale utilizzando la scansione I rispetto ai controlli.
Figura 1: mappe parametriche statistiche che dimostrano le differenze strutturali nella materia grigia nei pazienti con dolore cronico dovuto all'OA dell'anca primaria rispetto ai controlli e longitudinalmente rispetto a se stessi nel tempo. I cambiamenti significativi della materia grigia sono mostrati sovrapposti a colori, i dati della sezione trasversale sono rappresentati in rosso e i dati longitudinali in giallo. Piano assiale: il lato sinistro dell'immagine è il lato sinistro del cervello. in alto: aree di significativa diminuzione della materia grigia tra pazienti con dolore cronico dovuto a OA primaria dell'anca e soggetti di controllo non affetti. p <0.001 fondo non corretto: aumento della materia grigia in 20 pazienti senza dolore al terzo e quarto periodo di scansione dopo l'intervento di sostituzione totale dell'anca, rispetto alla prima (preoperatoria) e alla seconda (6-8 settimane dopo l'intervento). p <0.001 Grafici non corretti: stime del contrasto e intervallo di confidenza al 90%, effetti di interesse, unità arbitrarie. Asse x: contrasti per i 4 punti temporali, asse y: stima del contrasto a? 3, 50, 2 per ACC e stima del contrasto a 36, 39, 3 per insula.
Capovolgere i dati dei pazienti con OA dell'anca sinistra (n? =? 7) e confrontarli con controlli sani non ha modificato i risultati in modo significativo, ma per una diminuzione del talamo (x? =? 10, y? = ?? 20, z? =? 3, p <0.001, t? =? 3.44) e un aumento del cervelletto destro (x? =? 25, y? = ?? 37, z? = ?? 50, p <0.001, t? =? 5.12) che non ha raggiunto la significatività nei dati non ribaltati dei pazienti rispetto ai controlli.
Analisi longitudinale Nell'analisi longitudinale, è stato rilevato un aumento significativo (p <001 non corretto) della materia grigia confrontando la prima e la seconda scansione (dolore cronico / dolore post-operatorio) con la terza e la quarta scansione (senza dolore) nell'ACC, corteccia insulare, cervelletto e pars orbitalis nei pazienti con OA (Tabella 2 e Figura 1). La materia grigia è diminuita nel tempo (p <001 analisi dell'intero cervello non corretta) nella corteccia somatosensoriale secondaria, nell'ippocampo, nella corteccia midcingulata, nel talamo e nel nucleo caudato nei pazienti con OA (Figura 2).
Figura 2: a) Significativo aumento della materia grigia cerebrale a seguito di un'operazione riuscita Vista assiale di una significativa diminuzione della materia grigia nei pazienti con dolore cronico dovuto all'OA primaria dell'anca rispetto ai soggetti di controllo. p <0.001 non corretto (analisi della sezione trasversale), b) Aumento longitudinale della materia grigia nel tempo in giallo confrontando scansione I e II scansione III> scansione IV) in pazienti con OA. p <0.001 non corretto (analisi longitudinale). Il lato sinistro dell'immagine è il lato sinistro del cervello.
Il capovolgimento dei dati dei pazienti con OA dell'anca sinistra (n? =? 7) non ha modificato i risultati in modo significativo, ma per una diminuzione della materia grigia cerebrale nel giro di Heschl (x? = ?? 41, y? = ?? 21, z? =? 10, p <0.001, t? =? 3.69) e Precuneus (x? =? 15, y? = ?? 36, z? =? 3, p <0.001, t? =? 4.60) .
Confrontando la prima scansione (prechirurgia) con le scansioni 3 + 4 (postchirurgia), abbiamo riscontrato un aumento della materia grigia nella corteccia frontale e nella corteccia motoria (p <0.001 non corretto). Notiamo che questo contrasto è meno rigoroso poiché ora abbiamo meno scansioni per condizione (dolore vs non dolore). Quando abbassiamo la soglia ripetiamo ciò che abbiamo trovato usando il contrasto di 1 + 2 contro 3 + 4.
Cercando aree che aumentano in tutti gli intervalli di tempo, abbiamo trovato cambiamenti della materia grigia cerebrale nelle aree motorie (area 6) in pazienti con coxartrosi dopo la sostituzione totale dell'anca (scansione Idbm.neuro.uni-jena.de/vbm/) potremmo replicare questo risultato nella corteccia cingolata anteriore e media e in entrambe le insule anteriori.
Abbiamo calcolato le dimensioni dell'effetto e l'analisi trasversale (pazienti vs. controlli) ha prodotto un Cohen sd di 1.78751 nel voxel di picco dell'ACC (x? = ?? 12, y? =? 25, z? = ?? 16). Abbiamo anche calcolato Cohen sd per l'analisi longitudinale (scansione di contrasto 1 + 2 vs. scansione 3 + 4). Ciò ha portato a un Cohen sd di 1.1158 nell'ACC (x? = ?? 3, y? =? 50, z? =? 2). Per quanto riguarda l'insula (x? = ?? 33, y? =? 21, z? =? 13) e relativa allo stesso contrasto, Cohen sd è 1.0949. Inoltre, abbiamo calcolato la media dei valori voxel diversi da zero della mappa di Cohen sd all'interno della ROI (composta dalla divisione anteriore del giro cingolato e dalla corteccia subcallosa, derivata dall'Atlante strutturale corticale di Harvard-Oxford): 1.251223.
Insight di Dr. Alex Jimenez
I pazienti con dolore cronico possono sperimentare una varietà di problemi di salute nel tempo, a parte i loro sintomi già debilitanti. Per esempio, molti individui sperimenteranno problemi di sonno a causa del loro dolore, ma soprattutto, il dolore cronico può portare a vari problemi di salute mentale, tra cui ansia e depressione. Gli effetti che il dolore può avere sul cervello possono sembrare fin troppo schiaccianti, ma prove crescenti suggeriscono che questi cambiamenti cerebrali non sono permanenti e possono essere invertiti quando i pazienti con dolore cronico ricevono il trattamento adeguato per i loro problemi di salute di base. Secondo l'articolo, le anomalie della sostanza grigia riscontrate nel dolore cronico non riflettono il danno cerebrale, ma piuttosto sono una conseguenza reversibile che si normalizza quando il dolore viene adeguatamente trattato. Fortunatamente, sono disponibili una varietà di approcci terapeutici per alleviare i sintomi del dolore cronico e ripristinare la struttura e la funzione del cervello.
Discussione
Monitorando l'intera struttura cerebrale nel tempo, confermiamo ed espandiamo i nostri dati pilota pubblicati di recente [17]. Abbiamo trovato cambiamenti nella materia grigia cerebrale in pazienti con artrosi primaria dell'anca nello stato di dolore cronico, che si invertono in parte quando questi pazienti sono liberi dal dolore, dopo un intervento chirurgico endoprotesico dell'articolazione dell'anca. L'aumento parziale della materia grigia dopo l'intervento chirurgico è quasi nelle stesse aree in cui è stata osservata una diminuzione della materia grigia prima dell'intervento chirurgico. Capovolgere i dati dei pazienti con OA dell'anca sinistra (e quindi normalizzarsi per il lato del dolore) ha avuto solo un impatto minimo sui risultati, ma ha anche mostrato una diminuzione della materia grigia nel giro di Heschl e nel Precuneus che non possiamo facilmente spiegare e poiché non esistono ipotesi a priori, guardatelo con grande cautela. Tuttavia, la differenza osservata tra pazienti e controlli sani alla scansione I era ancora osservabile nell'analisi della sezione trasversale alla scansione IV. L'aumento relativo della materia grigia nel tempo è quindi sottile, cioè non sufficientemente distinto da avere un effetto sull'analisi della sezione trasversale, una scoperta che è già stata dimostrata negli studi che indagano la plasticità dipendente dall'esperienza [30], [31]. Notiamo che il fatto che mostriamo come reversibili alcune parti dei cambiamenti cerebrali dovuti al dolore cronico non esclude che alcune altre parti di questi cambiamenti siano irreversibili.
È interessante notare che la diminuzione della sostanza grigia nell'ACC in pazienti con dolore cronico prima dell'intervento sembra continuare 6 settimane dopo l'intervento chirurgico (scansione II) e aumenta solo verso la scansione III e IV, probabilmente a causa di dolore post-operatorio o diminuzione del motore funzione. Questo è in linea con i dati comportamentali del punteggio di mobilità fisica incluso nel NHP, che dopo l'intervento non mostrava alcun cambiamento significativo al punto di tempo II, ma aumentava significativamente verso la scansione III e IV. Di nota, i nostri pazienti non hanno riportato dolore all'anca dopo l'intervento chirurgico, ma hanno sperimentato dolore post-operatorio nei muscoli circostanti e nella pelle che è stato percepito in modo molto diverso dai pazienti. Tuttavia, poiché i pazienti riportavano ancora dolore alla scansione II, abbiamo anche confrontato la prima scansione (pre-intervento) con le scansioni III + IV (post-chirurgia), rivelando un aumento della materia grigia nella corteccia frontale e nella corteccia motoria. Notiamo che questo contrasto è meno rigoroso a causa di meno scansioni per condizione (dolore vs. non dolore). Quando abbassiamo la soglia ripetiamo ciò che abbiamo trovato usando il contrasto di I + II vs. III + IV.
I nostri dati suggeriscono fortemente che le alterazioni della materia grigia nei pazienti con dolore cronico, che di solito si trovano nelle aree coinvolte nell'elaborazione nocicettiva sopraspinale [4], non sono né dovute ad atrofia neuronale né danno cerebrale. Il fatto che questi cambiamenti osservati nello stato di dolore cronico non si annullino completamente potrebbe essere spiegato con il periodo di osservazione relativamente breve (un anno dopo l'operazione contro una media di sette anni di dolore cronico prima dell'intervento). I cambiamenti cerebrali neuroplastici che possono essersi sviluppati nel corso di diversi anni (come conseguenza del costante input nocicettivo) richiedono probabilmente più tempo per invertirsi completamente. Un'altra possibilità per cui l'aumento della materia grigia può essere rilevato solo nei dati longitudinali ma non nei dati trasversali (cioè tra coorti al punto temporale IV) è che il numero di pazienti (n? =? 20) è troppo piccolo. Va sottolineato che la varianza tra i cervelli di più individui è piuttosto ampia e che i dati longitudinali hanno il vantaggio che la varianza è relativamente piccola poiché gli stessi cervelli vengono scansionati più volte. Di conseguenza, i cambiamenti sottili saranno rilevabili solo nei dati longitudinali [30], [31], [32]. Ovviamente non possiamo escludere che questi cambiamenti siano almeno in parte irreversibili sebbene ciò sia improbabile, dati i risultati della plasticità strutturale specifica dell'esercizio e della riorganizzazione [4], [12], [30], [33], [34]. Per rispondere a questa domanda, studi futuri devono indagare ripetutamente sui pazienti per periodi di tempo più lunghi, possibilmente anni.
Notiamo che possiamo solo trarre conclusioni limitate riguardo alla dinamica dei cambiamenti morfologici del cervello nel tempo. Il motivo è che quando abbiamo progettato questo studio in 2007 e scansionato in 2008 e 2009, non era noto se si sarebbero verificati cambiamenti strutturali e, per motivi di fattibilità, abbiamo scelto le date e gli intervalli di scansione come descritto qui. Si potrebbe obiettare che la materia grigia cambia nel tempo, che descriviamo per il gruppo di pazienti, potrebbe anche essersi verificata nel gruppo di controllo (effetto del tempo). Tuttavia, eventuali modifiche dovute all'invecchiamento dovrebbero essere una diminuzione del volume. Data la nostra ipotesi a priori, basata su studi indipendenti di 9 e coorti che mostrano diminuzioni della materia grigia nei pazienti con dolore cronico [7], [8], [9], [15], [24], [25], [26], [27], [28], ci siamo concentrati sugli aumenti regionali nel tempo e quindi riteniamo che non si tratti di un semplice effetto del tempo. Da notare, non possiamo escludere che la materia grigia diminuisca nel tempo che abbiamo riscontrato nel nostro gruppo di pazienti a causa di un effetto del tempo, poiché non abbiamo scansionato il nostro gruppo di controllo nello stesso intervallo di tempo. Alla luce dei risultati, gli studi futuri dovrebbero mirare a intervalli di tempo sempre più brevi, dato che i cambiamenti morfometrici dipendenti dall'esercizio possono verificarsi rapidamente come dopo 1 settimana [32], [33].
Oltre all'impatto dell'aspetto nocicettivo del dolore sulla materia grigia del cervello [17], [34] abbiamo osservato che i cambiamenti nella funzione motoria probabilmente contribuiscono anche ai cambiamenti strutturali. Abbiamo trovato aree motore e premotorie (area 6) per aumentare su tutti gli intervalli di tempo (Figura 3). Intuitivamente ciò potrebbe essere dovuto al miglioramento della funzione motoria nel tempo poiché i pazienti non erano più limitati nel vivere una vita normale. In particolare, non ci siamo concentrati sulla funzione motoria, ma su un miglioramento nell'esperienza del dolore, data la nostra ricerca originale per indagare se la ben nota riduzione della materia grigia cerebrale nei pazienti con dolore cronico è in linea di principio reversibile. Di conseguenza, non abbiamo utilizzato strumenti specifici per studiare la funzione motoria. Tuttavia, la riorganizzazione della corteccia motoria (funzionale) nei pazienti con sindromi dolorose è ben documentata [35], [36], [37], [38]. Inoltre, la corteccia motoria è un bersaglio negli approcci terapeutici in pazienti con dolore cronico clinicamente intrattabile mediante stimolazione cerebrale diretta [39], [40], stimolazione transcranica in corrente continua [41] e stimolazione magnetica transcranica ripetitiva [42], [43]. I meccanismi esatti di tale modulazione (facilitazione contro inibizione o semplicemente interferenza nelle reti correlate al dolore) non sono ancora stati chiariti [40]. Uno studio recente ha dimostrato che un'esperienza motoria specifica può alterare la struttura del cervello [13]. La sincatogenesi, la riorganizzazione delle rappresentazioni del movimento e l'angiogenesi nella corteccia motoria possono verificarsi con richieste speciali di un compito motorio. Tsao et al. ha mostrato riorganizzazione nella corteccia motoria di pazienti con lombalgia cronica che sembrano essere specifici per il dolore alla schiena [44] e Puri et al. osservato una riduzione della materia grigia della zona motoria supplementare sinistra in soggetti affetti da fibromialgia [45]. Il nostro studio non è stato progettato per districare i diversi fattori che possono cambiare il cervello nel dolore cronico, ma interpretiamo i nostri dati riguardanti i cambiamenti della materia grigia che non rispecchiano esclusivamente le conseguenze dell'input nocicettivo costante. Infatti, un recente studio in pazienti con dolore neuropatico ha evidenziato anomalie nelle regioni del cervello che comprendono la percezione emotiva, autonomica e del dolore, implicando che giocano un ruolo fondamentale nel quadro clinico globale del dolore cronico [28].
Figura 3: Mappe parametriche statistiche che dimostrano un aumento significativo della materia grigia cerebrale nelle aree motorie (area 6) in pazienti con coxartrosi prima rispetto a dopo THR (analisi longitudinale, scansione I Stime del contrasto in x? =? 19, y? = ?? 12, z? =? 70.
Due recenti studi pilota si sono concentrati sulla terapia di sostituzione dell'anca in pazienti con osteoartrite, l'unica sindrome da dolore cronico che è principalmente curabile con la sostituzione totale dell'anca [17], [46] e questi dati sono affiancati da uno studio molto recente in pazienti cronici di lombalgia [ 47]. Questi studi devono essere visti alla luce di diversi studi longitudinali che studiano la plasticità neuronale dipendente dall'esperienza a livello strutturale [30], [31] e un recente studio sui cambiamenti strutturali del cervello in volontari sani che sperimentano ripetute stimolazioni dolorose [34] . Il messaggio chiave di tutti questi studi è che la principale differenza nella struttura del cervello tra pazienti con dolore e controlli può diminuire quando il dolore è guarito. Tuttavia, si deve tener conto del fatto che semplicemente non è chiaro se i cambiamenti nei pazienti con dolore cronico siano dovuti unicamente a input nocicettivi oa causa delle conseguenze del dolore o di entrambi. È più che probabile che i cambiamenti comportamentali, come la privazione o l'aumento dei contatti sociali, l'agilità, l'allenamento fisico e le modifiche allo stile di vita siano sufficienti per modellare il cervello [6], [12], [28], [48]. Particolarmente la depressione come co-morbilità o conseguenza del dolore è un candidato chiave per spiegare le differenze tra pazienti e controlli. Un piccolo gruppo di nostri pazienti con OA ha mostrato sintomi depressivi lievi o moderati che sono cambiati nel tempo. Non abbiamo trovato le alterazioni strutturali di covary in modo significativo con il punteggio BDI, ma sorge la domanda su quanti altri cambiamenti comportamentali dovuti all'assenza di dolore e miglioramento motorio possono contribuire ai risultati e fino a che punto lo fanno. Questi cambiamenti comportamentali possono eventualmente influenzare una diminuzione della materia grigia nel dolore cronico e un aumento della sostanza grigia quando il dolore scompare.
Un altro fattore importante che può influenzare la nostra interpretazione dei risultati è il fatto che quasi tutti i pazienti con dolore cronico hanno assunto farmaci contro il dolore, che hanno interrotto quando erano senza dolore. Si potrebbe argomentare che i FANS come il diclofenac o l'ibuprofene hanno alcuni effetti sui sistemi neurali e lo stesso vale per gli oppioidi, gli antiepilettici e gli antidepressivi, farmaci che sono frequentemente usati nella terapia del dolore cronico. L'impatto di antidolorifici e altri farmaci sui risultati morfometrici potrebbe essere importante (48). Nessuno studio ha finora mostrato effetti del farmaco antidolorifico sulla morfologia del cervello, ma diversi articoli hanno scoperto che i cambiamenti nella struttura cerebrale nei pazienti con dolore cronico non sono spiegati solo dall'inattività associata al dolore [15], né dai farmaci antidolorifici [7], [9], [49]. Tuttavia, mancano studi specifici. Ulteriori ricerche dovrebbero focalizzare i cambiamenti dipendenti dall'esperienza nella plasticità corticale, che possono avere vaste implicazioni cliniche per il trattamento del dolore cronico.
Abbiamo anche riscontrato diminuzioni della materia grigia nell'analisi longitudinale, probabilmente dovute a processi di riorganizzazione che accompagnano i cambiamenti nella funzione motoria e nella percezione del dolore. Ci sono poche informazioni disponibili sui cambiamenti longitudinali nella materia grigia del cervello in condizioni di dolore, per questo motivo non abbiamo alcuna ipotesi per una diminuzione della sostanza grigia in queste aree dopo l'operazione. Teutsch et al. [25] ha riscontrato un aumento della materia grigia cerebrale nella corteccia somatosensoriale e midcingolata in volontari sani che hanno sperimentato una stimolazione dolorosa in un protocollo giornaliero per otto giorni consecutivi. Il reperto di sostanza grigia aumenta a seguito di input nocicettivo sperimentale sovrapposto anatomicamente in una certa misura alla diminuzione della materia grigia cerebrale in questo studio in pazienti che sono stati curati da un dolore cronico di lunga durata. Ciò implica che l'input nocicettivo in volontari sani porta all'esercizio di cambiamenti strutturali dipendenti, come nei pazienti con dolore cronico, e che questi cambiamenti si invertono in volontari sani quando l'input nocicettivo si arresta. Di conseguenza, la diminuzione della materia grigia in queste aree osservate in pazienti con OA potrebbe essere interpretata in modo da seguire lo stesso processo fondamentale: cambiamenti dipendenti dall'esercizio cambiamenti cerebrali [50]. Come procedura non invasiva, MR Morphometry è lo strumento ideale per la ricerca dei substrati morfologici delle malattie, approfondendo la nostra comprensione della relazione tra struttura e funzione del cervello e persino per monitorare gli interventi terapeutici. Una delle grandi sfide in futuro è quella di adattare questo potente strumento per le prove multicentriche e terapeutiche del dolore cronico.
Limitazioni di questo studio
Sebbene questo studio sia un'estensione del nostro precedente studio che estende i dati di follow-up a 12 mesi e indaga su più pazienti, il nostro principio che trova che i cambiamenti morfometrici del cervello nel dolore cronico sono reversibili è piuttosto sottile. Le dimensioni degli effetti sono piccole (vedi sopra) e gli effetti sono in parte guidati da un'ulteriore riduzione del volume di materia grigia cerebrale regionale al punto temporale della scansione 2. Quando escludiamo i dati dalla scansione 2 (subito dopo l'operazione) solo significativi gli aumenti della materia grigia cerebrale per la corteccia motoria e la corteccia frontale sopravvivono a una soglia di p <0.001 non corretta (Tabella 3).
Conclusione
Non è possibile distinguere fino a che punto le alterazioni strutturali che abbiamo osservato siano dovute a cambiamenti nell'input nocicettivo, cambiamenti nella funzione motoria o consumo di farmaci o cambiamenti nel benessere in quanto tali. Mascherare i contrasti di gruppo della prima e dell'ultima scansione mostrava differenze molto minori del previsto. Presumibilmente, le alterazioni cerebrali dovute a dolore cronico con tutte le conseguenze si stanno sviluppando in un periodo piuttosto lungo e potrebbe anche aver bisogno di tempo per tornare indietro. Tuttavia, questi risultati rivelano processi di riorganizzazione, suggerendo fortemente che l'input nocicettivo cronico e la compromissione motoria in questi pazienti portano ad un alterato processo nelle regioni corticali e conseguentemente a cambiamenti cerebrali strutturali che sono in linea di principio reversibili.
Ringraziamenti
Ringraziamo tutti i volontari per la partecipazione a questo studio e al gruppo di fisica e metodi presso NeuroImage Nord ad Amburgo. Lo studio ha ottenuto l'approvazione etica da parte del comitato etico locale e il consenso informato scritto è stato ottenuto da tutti i partecipanti allo studio prima dell'esame.
Dichiarazione di finanziamento
Questo lavoro è stato supportato da sovvenzioni della DFG (Fondazione tedesca per la ricerca) (MA 1862 / 2-3) e BMBF (Ministero federale dell'Istruzione e della ricerca) (371 57 01 e NeuroImage Nord). I finanziatori non hanno avuto alcun ruolo nella progettazione dello studio, nella raccolta e analisi dei dati, nella decisione di pubblicare o nella preparazione del manoscritto.
Il sistema endocannabinoide: il sistema essenziale di cui non hai mai sentito parlare
Nel caso in cui non hai sentito parlare del sistema endocannabinoide, o ECS, non c'è bisogno di sentirsi in imbarazzo. Di nuovo negli 1960, gli investigatori che si interessarono alla bioattività della Cannabis alla fine isolarono molte delle sue sostanze chimiche attive. Ci sono voluti altri anni 30, tuttavia, per i ricercatori che studiano modelli animali per trovare un recettore per questi prodotti chimici ECS nel cervello dei roditori, una scoperta che ha aperto un intero mondo di indagine sull'esistenza dei recettori ECS e qual è il loro scopo fisiologico.
Ora sappiamo che la maggior parte degli animali, dal pesce agli uccelli ai mammiferi, possiede un endocannabinoide e sappiamo che gli esseri umani non solo producono i loro cannabinoidi che interagiscono con questo particolare sistema, ma produciamo anche altri composti che interagiscono con gli ECS, quelli di che sono osservati in molte piante e cibi diversi, ben oltre le specie di Cannabis.
Come sistema del corpo umano, l'ECS non è una piattaforma strutturale isolata come il sistema nervoso o il sistema cardiovascolare. Invece, l'ECS è un insieme di recettori ampiamente distribuiti in tutto il corpo che sono attivati attraverso una serie di ligandi che collettivamente conosciamo come endocannabinoidi o cannabinoidi endogeni. Entrambi i recettori verificati sono chiamati solo CB1 e CB2, sebbene ve ne siano altri proposti. Anche i canali PPAR e TRP mediano alcune funzioni. Allo stesso modo, troverete solo due endocannabinoidi ben documentati: anadamide e 2-arachidonoil glicerolo o 2-AG.
Inoltre, fondamentali per il sistema endocannabinoide sono gli enzimi che sintetizzano e abbattono gli endocannabinoidi. Si ritiene che gli endocannabinoidi siano sintetizzati in una base necessaria. Gli enzimi primari coinvolti sono diacilglicerolo-lipasi e N-acil-fosfatidiletanolammina-fosfolipasi D, che sintetizzano rispettivamente 2-AG e anandamide. I due principali enzimi degradanti sono l'acido grasso ammide idrolasi, o FAAH, che scompone l'anandamide e il monoacilglicerolo lipasi, o MAGL, che scompone 2-AG. La regolazione di questi due enzimi può aumentare o diminuire la modulazione dell'ECS.
Qual è la funzione di ECS?
L'ECS è il principale sistema di regolazione omeostatica del corpo. Può essere facilmente visto come il sistema adattogeno interno del corpo, lavorando sempre per mantenere l'equilibrio di una varietà di funzioni. Gli endocannabinoidi funzionano in generale come neuromodulatori e, come tali, regolano un'ampia gamma di processi corporei, dalla fertilità al dolore. Alcune delle funzioni più note di ECS sono le seguenti:
Sistema Nervoso
Dal sistema nervoso centrale o dal SNC, la stimolazione generale dei recettori CB1 inibisce il rilascio di glutammato e GABA. Nel sistema nervoso centrale, l'ECS svolge un ruolo nella formazione e apprendimento della memoria, promuove la neurogenesi nell'ippocampo, regola anche l'eccitabilità neuronale. L'ECS svolge anche un ruolo nel modo in cui il cervello reagisce a lesioni e infiammazioni. Dal midollo spinale, l'ECS modula la segnalazione del dolore e aumenta l'analgesia naturale. Nel sistema nervoso periferico, nel quale i recettori CB2 controllano, l'ECS agisce principalmente nel sistema nervoso simpatico per regolare le funzioni dei tratti intestinali, urinari e riproduttivi.
Stress e umore
L'ECS ha molteplici impatti sulle reazioni di stress e sulla regolazione emotiva, come l'inizio di questa risposta corporea allo stress acuto e l'adattamento nel tempo a più emozioni a lungo termine, come la paura e l'ansia. Un sano sistema endocannabinoide funzionante è fondamentale per il modo in cui gli umani modulano tra un soddisfacente grado di eccitazione rispetto a un livello eccessivo e sgradevole. L'ECS svolge anche un ruolo nella formazione della memoria e forse soprattutto nel modo in cui il cervello imprime i ricordi a stress o lesioni. Poiché l'ECS modula il rilascio di dopamina, noradrenalina, serotonina e cortisolo, può anche influenzare ampiamente la risposta emotiva e i comportamenti.
Apparato digerente
Il tubo digerente è popolato con i recettori CB1 e CB2 che regolano diversi aspetti importanti della salute GI. Si è pensato che l'ECS potrebbe essere il "collegamento mancante" nel descrivere il legame gastro-cervello-immuno che svolge un ruolo significativo nella salute funzionale del tratto digestivo. L'ECS è un regolatore dell'immunità intestinale, forse limitando il sistema immunitario a distruggere la flora sana e anche attraverso la modulazione della segnalazione delle citochine. L'ECS modula la risposta infiammatoria naturale nel tratto digestivo, che ha importanti implicazioni per una vasta gamma di problemi di salute. Anche la motilità gastrointestinale e generale sembra essere parzialmente governata dall'ECS.
Appetito e metabolismo
L'ECS, in particolare i recettori CB1, svolge un ruolo nell'appetito, nel metabolismo e nella regolazione del grasso corporeo. La stimolazione dei recettori CB1 aumenta il comportamento alla ricerca di cibo, migliora la consapevolezza dell'olfatto, regola anche il bilancio energetico. Sia gli animali che gli esseri umani che sono in sovrappeso hanno una disregolazione ECS che può portare questo sistema a diventare iperattivo, il che contribuisce sia alla sovralimentazione che alla riduzione del dispendio energetico. È stato dimostrato che i livelli circolanti di anandamide e 2-AG sono elevati nell'obesità, che potrebbe essere in parte dovuta alla diminuzione della produzione dell'enzima di degradazione FAAH.
Salute immunitaria e risposta infiammatoria
Le cellule e gli organi del sistema immunitario sono ricchi di recettori endocannabinoidi. I recettori dei cannabinoidi sono espressi nella ghiandola del timo, nella milza, nelle tonsille e nel midollo osseo, nonché nei linfociti T e B, nei macrofagi, nei mastociti, nei neutrofili e nelle cellule natural killer. L'ECS è considerato il principale motore dell'equilibrio e dell'omeostasi del sistema immunitario. Sebbene non tutte le funzioni dell'ECS del sistema immunitario siano comprese, l'ECS sembra regolare la produzione di citochine e anche avere un ruolo nel prevenire l'iperattività nel sistema immunitario. L'infiammazione è una parte naturale della risposta immunitaria e svolge un ruolo molto normale negli insulti acuti all'organismo, tra cui lesioni e malattie; tuttavia, quando non viene tenuto sotto controllo può diventare cronico e contribuire a una cascata di problemi di salute avversi, come il dolore cronico. Mantenendo sotto controllo la risposta immunitaria, l'ECS aiuta a mantenere una risposta infiammatoria più equilibrata attraverso il corpo.
Altre aree di salute regolate dalla ECS:
La salute delle ossa
Fertilità
Salute della pelle
Salute arteriosa e respiratoria
Sonno e ritmo circadiano
Come sostenere al meglio un ECS in buona salute è una domanda a cui molti ricercatori stanno ora cercando di rispondere. Restate sintonizzati per ulteriori informazioni su questo argomento emergente.
In conclusione,Il dolore cronico è stato associato a cambiamenti cerebrali, inclusa la riduzione della materia grigia. Tuttavia, l'articolo sopra ha dimostrato che il dolore cronico può alterare la struttura e la funzione complessiva del cervello. Sebbene il dolore cronico possa portare a questi, tra gli altri problemi di salute, il trattamento adeguato dei sintomi sottostanti del paziente può invertire i cambiamenti cerebrali e regolare la materia grigia. Inoltre, sono emersi sempre più studi di ricerca sull'importanza del sistema endocannabinoide e sulla sua funzione nel controllo e nella gestione del dolore cronico e di altri problemi di salute. Informazioni referenziate dal National Center for Biotechnology Information (NCBI) . L'ambito delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
1. Woolf CJ, Salter MW (2000) Plasticità neuronale: aumento del guadagno di dolore. Scienzeï ¿½288: 1765 1769.[PubMed]
2. Flor H, Nikolajsen L, Staehelin Jensen T (2006) Dolore agli arti fantasma: un caso di plasticità maladattiva del SNC?ï ¿½Nat Rev Neurosciï ¿½7: 873 881. [PubMed]
3. Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC, et al. (2009) Cambiamenti anatomici nella corteccia motoria umana e nei percorsi motori dopo la completa lesione del midollo spinale toracico. Cereb Cortexï ¿½19: 224 232. [PubMed]
4. Maggio A (2008) Il dolore cronico può cambiare la struttura del cervello. Doloreï ¿½137: 7 15. [PubMed]
5. May A (2009) Morphing voxel: l'hype intorno all'immagine strutturale dei pazienti con cefalea. Cervello.[PubMed]
6. Apkarian AV, Baliki MN, Geha PY (2009) Verso una teoria del dolore cronico. Prog Neurobiolï ¿½87: 81 97. [Articolo gratuito di PMC]ï ¿½[PubMed]
7. Apkarian AV, Sosa Y, Sonty S, Levy RM, Harden RN, et al. (2004) Il mal di schiena cronico è associato a una diminuzione della densità della materia grigia prefrontale e talamica. J Neurosciï ¿½24: 10410 10415. [PubMed]
8. Rocca MA, Ceccarelli A, Falini A, Colombo B, Tortorella P, et al. (2006) Cambiamenti della materia grigia cerebrale nei pazienti con emicrania con lesioni visibili T2: uno studio RMN 3-T. Colpoï ¿½37: 1765 1770. [PubMed]
9. Kuchinad A, Schweinhardt P, Seminowicz DA, Wood PB, Chizh BA, et al. (2007) Accelerazione della perdita di materia grigia cerebrale nei pazienti con fibromialgia: invecchiamento precoce del cervello?ï ¿½J Neurosciï ¿½27: 4004 4007.[PubMed]
10. Tracey I, Bushnell MC (2009) Come gli studi di neuroimaging ci hanno sfidato a ripensare: il dolore cronico è una malattia?ï ¿½J Painï ¿½10: 1113 1120. [PubMed]
11. Franke K, Ziegler G, Kloppel S, Gaser C (2010) Stima dell'età dei soggetti sani dalle scansioni MRI pesate con T1 utilizzando i metodi del kernel: esplorazione dell'influenza di vari parametri. Neuroimageï ¿½50: 883 892. [PubMed]
12. Draganski B, maggio A (2008) Cambiamenti strutturali indotti dall'addestramento nel cervello umano adulto. Behav Brain Resï ¿½192: 137 142. [PubMed]
13. Adkins DL, Boychuk J, Remple MS, Kleim JA (2006) L'allenamento motorio induce modelli di plasticità specifici dell'esperienza attraverso la corteccia motoria e il midollo spinale. J Appl Physiolï ¿½101: 1776 1782. [PubMed]
14. Duerden EG, Laverdure-Dupont D (2008) La pratica fa la corteccia. J Neurosciï ¿½28: 8655 8657. [PubMed]
15. Draganski B, Moser T, Lummel N, Ganssbauer S, Bogdahn U, et al. (2006) Diminuzione della materia grigia talamica dopo l'amputazione degli arti. Neuroimageï ¿½31: 951 957. [PubMed]
16. Nikolajsen L, Brandsborg B, Lucht U, Jensen TS, Kehlet H (2006) Dolore cronico dopo artroplastica totale d'anca: uno studio nazionale sul questionario. Acta Anaesthesiol Scandï ¿½50: 495 500. [PubMed]
17. Rodriguez-Raecke R, Niemeier A, Ihle K, Ruether W, May A (2009) La diminuzione della materia grigia cerebrale nel dolore cronico è la conseguenza e non la causa del dolore. J Neurosciï ¿½29: 13746 13750. [PubMed]
18. Beck AT, Ward CH, Mendelson M, Mock J, Erbaugh J (1961) Un inventario per misurare la depressione. Arch Gen Psychiatryï ¿½4: 561 571. [PubMed]
19. Franke G (2002) Die Symptom-Checkliste nach LR Derogatis - Manual. G ttingen Beltz Test Verlag.
20. Geissner E (1995) The Pain Perception Scale, una scala differenziata e sensibile al cambiamento per la valutazione del dolore cronico e acuto. Riabilitazione (Stuttg) 34: XXXV XLIII. [PubMed]
21. Bullinger M, Kirchberger I (1998) SF-36 - Fragebogen zum Gesundheitszustand. Hand-anweisung. G ttingen: Hogrefe.
22. Ashburner J, Friston KJ (2000) Morfometria basata su voxel: i metodi. Neuroimageï ¿½11: 805 821.[PubMed]
23. Buon CD, Johnsrude IS, Ashburner J, Henson RN, Friston KJ, et al. (2001) Uno studio morfometrico basato sul voxel dell'invecchiamento nei cervelli umani adulti 465 normali. Neuroimageï ¿½14: 21 36. [PubMed]
24. Baliki MN, Chialvo DR, Geha PY, Levy RM, Harden RN, et al. (2006) Dolore cronico e cervello emotivo: attività cerebrale specifica associata a fluttuazioni spontanee dell'intensità del mal di schiena cronico. J Neurosciï ¿½26: 12165 12173. [Articolo gratuito di PMC]ï ¿½[PubMed]
25. Lutz J, Jager L, de Quervain D, Krauseneck T, Padberg F, et al. (2008) Anomalie della materia bianca e grigia nel cervello dei pazienti con fibromialgia: studio di tensore della diffusione e volumetrico. artrite Rheumï ¿½58: 3960 3969. [PubMed]
26. Wrigley PJ, Gustin SM, Macey PM, Nash PG, Gandevia SC, et al. (2008) Cambiamenti anatomici nella corteccia motoria umana e nei percorsi motori dopo la lesione completa del midollo spinale toracico. Cereb Cortex19: 224 232. [PubMed]
27. Schmidt-Wilcke T, Hierlmeier S, Leinisch E (2010) Altered Regional Brain Morphology in Patients With Chronic Facial Pain. Mal di testa[PubMed]
28. Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB, et al. (2008) Il cervello nel dolore cronico CRPS: anormali interazioni grigio-bianco nelle regioni emotive e autonome. Neuronï ¿½60: 570 581. [Articolo gratuito di PMC]ï ¿½[PubMed]
29. Braciere J, Roberts J, Deverill M (2002) La stima di una misura di salute basata sulle preferenze dall'SF-36. J Health Econï ¿½21: 271 292. [PubMed]
30. Draganski B, Gaser C, Busch V, Schuierer G, Bogdahn U, et al. (2004) Neuroplasticità: cambiamenti nella materia grigia indotti dall'allenamento. Naturaï ¿½427: 311 312. [PubMed]
31. Boyke J, Driemeyer J, Gaser C, Buchel C, May A (2008) Cambiamenti nella struttura cerebrale indotti dall'addestramento negli anziani. J Neurosciï ¿½28: 7031 7035. [PubMed]
32. Driemeyer J, Boyke J, Gaser C, Buchel C, May A (2008) Cambiamenti nella materia grigia indotti dall'apprendimento rivisitato. PLoS ONEï ¿½3: e2669. [Articolo gratuito di PMC]ï ¿½[PubMed]
33. May A, Hajak G, Ganssbauer S, Steffens T, Langguth B, et al. (2007) Alterazioni strutturali del cervello dopo 5 giorni di intervento: aspetti dinamici della neuroplasticità. Cereb Cortexï ¿½17: 205 210. [PubMed]
34. Teutsch S, Herken W, Bingel U, Schoell E, May A (2008) Cambiamenti nella materia grigia del cervello a causa della stimolazione dolorosa ripetitiva. Neuroimageï ¿½42: 845 849. [PubMed]
35. Flor H, Braun C, Elbert T, Birbaumer N (1997) Ampia riorganizzazione della corteccia somatosensoriale primaria nei pazienti cronici di mal di schiena. Neurosci Lettï ¿½224: 5 8. [PubMed]
36. Flor H, Denke C, Schaefer M, Grusser S (2001) Effetto dell'allenamento sulla discriminazione sensoriale sulla riorganizzazione corticale e sul dolore degli arti fantasma. Lancettaï ¿½357: 1763 1764. [PubMed]
37. Swart CM, Stins JF, Beek PJ (2009) Cambiamenti corticali nella sindrome da dolore regionale complesso (CRPS). Eur J Painï ¿½13: 902 907. [PubMed]
38. Maihofner C, Baron R, DeCol R, Binder A, Birklein F, et al. (2007) Il sistema motorio mostra cambiamenti adattativi nella sindrome dolorosa regionale complessa. Cervelloï ¿½130: 2671 2687. [PubMed]
39. Fontaine D, Hamani C, Lozano A (2009) Efficacia e sicurezza della stimolazione della corteccia motoria per il dolore neuropatico cronico: revisione critica della letteratura. J Neurosurgï ¿½110: 251 256. [PubMed]
40. Levy R, Deer TR, Henderson J (2010) Neurostimolazione intracranica per il controllo del dolore: una revisione. Medico del doloreï ¿½13: 157 165. [PubMed]
41. Antal A, Brepohl N, Poreisz C, Boros K, Csifcsak G, et al. (2008) La stimolazione della corrente continua transcranica sulla corteccia somatosensoriale riduce la percezione del dolore acuto indotta sperimentalmente. Clin J Pain24: 56 63. [PubMed]
42. Teepker M, Hotzel J, Timmesfeld N, Reis J, Mylius V, et al. (2010) RTMS a bassa frequenza del vertice nel trattamento profilattico dell'emicrania. Cefaleeï ¿½30: 137 144. [PubMed]
43. O'Connell N, Wand B, Marston L, Spencer S, Desouza L (2010) Tecniche di stimolazione cerebrale non invasiva per dolore cronico. Una relazione di una revisione sistematica di Cochrane e una meta-analisi. Eur J Phys Rehabil Medï ¿½47: 309 326. [PubMed]
44. Tsao H, Galea MP, Hodges PW (2008) La riorganizzazione della corteccia motoria è associata a deficit di controllo posturale nella lombalgia ricorrente. Cervelloï ¿½131: 2161 2171. [PubMed]
45. Puri BK, Agour M, Gunatilake KD, Fernando KA, Gurusinghe AI, et al. (2010) Riduzione della materia grigia della zona motoria supplementare sinistra in soggetti adulti affetti da fibromialgia femminile con affaticamento marcato e senza disturbi affettivi: studio di morfometria basato su voxel basato sulla risonanza magnetica 3-T pilotato. J Int Med Resï ¿½38: 1468 1472. [PubMed]
46. Gwilym SE, Fillipini N, Douaud G, Carr AJ, Tracey I (2010) L'atrofia talamica associata all'osteoartrosi dolorosa dell'anca è reversibile dopo l'artroprotesi; uno studio morfometrico longitudinale basato su voxel. Artrite reumata[PubMed]
47. Seminowicz DA, Wideman TH, Naso L, Hatami-Khoroushahi Z, Fallatah S, et al. (2011) L'efficace trattamento della lombalgia cronica negli umani inverte anomalie anatomiche e funzionali del cervello. J Neurosci31: 7540 7550. [PubMed]
48. May A, Gaser C (2006) Morfometria basata sulla risonanza magnetica: una finestra sulla plasticità strutturale del cervello. Curr Opin Neurolï ¿½19: 407 411. [PubMed]
49. Schmidt-Wilcke T, Leinisch E, Straube A, Kampfe N, Draganski B, et al. (2005) La sostanza grigia diminuisce nei pazienti con cefalea tipo tensione cronica. Neurologiaï ¿½65: 1483 1486. [PubMed]
50. Maggio A (2009) Morphing voxels: il clamore intorno all'imaging strutturale dei mal di testa. Cervello 132 (Pt6): 1419 1425. [PubMed]
Biochimica del dolore:Tutte le sindromi dolorose hanno un profilo infiammatorio. Un profilo infiammatorio può variare da persona a persona e può anche variare in una persona in momenti diversi. Il trattamento delle sindromi dolorose consiste nel comprendere questo profilo infiammatorio. Le sindromi dolorose sono trattate da un punto di vista medico, chirurgico o entrambi. L'obiettivo è inibire / sopprimere la produzione di mediatori infiammatori. E un risultato positivo è quello che si traduce in meno infiammazione e ovviamente meno dolore.
Biochimica del dolore
Obiettivi:
Chi sono i giocatori chiave
Quali sono i meccanismi biochimici?
Quali sono le conseguenze?
Recensione di infiammazione:
Giocatori chiave
Perché mi fa male la spalla? Una rassegna delle basi neuroanatomiche e biochimiche del dolore alla spalla
ABSTRACT
Se un paziente chiede `` perché mi fa male la spalla? '', La conversazione si sposterà rapidamente sulla teoria scientifica e talvolta su congetture prive di fondamento. Spesso, il clinico diventa consapevole dei limiti delle basi scientifiche della loro spiegazione, dimostrando l'incompletezza della nostra comprensione della natura del dolore alla spalla. Questa revisione adotta un approccio sistematico per aiutare a rispondere a domande fondamentali relative al dolore alla spalla, al fine di fornire approfondimenti sulla ricerca futura e nuovi metodi per il trattamento del dolore alla spalla. Esploreremo i ruoli di (1) i recettori periferici, (2) l'elaborazione del dolore periferico o nocicezione , (3) il midollo spinale, (4) il cervello, (5) la posizione dei recettori nella spalla e (6 ) l'anatomia neurale della spalla. Consideriamo anche come questi fattori possano contribuire alla variabilità nella presentazione clinica, nella diagnosi e nel trattamento del dolore alla spalla. In questo modo miriamo a fornire una panoramica delle parti componenti del sistema di rilevamento del dolore periferico e dei meccanismi centrali di elaborazione del dolore nel dolore alla spalla che interagiscono per produrre dolore clinico.
INTRODUZIONE: UNA STORIA MOLTO BREVE DI SCIENZA DEL DOLORE ESSENZIALE PER I CLINICI
La natura del dolore, in generale, è stata oggetto di molte controversie nel secolo scorso. Nel 17 ° secolo la teoria di Descartes1 propose che l'intensità del dolore fosse direttamente correlata alla quantità di lesione tissutale associata e che il dolore fosse elaborato in un percorso distinto. Molte teorie precedenti si basavano su questa cosiddetta filosofia cartesiana "dualista", vedendo il dolore come la conseguenza della stimolazione di un recettore del dolore periferico "specifico" nel cervello. Nel 20 ° secolo seguì una battaglia scientifica tra due teorie opposte, vale a dire la teoria della specificità e la teoria dei modelli. La `` teoria della specificità '' cartesiana vedeva il dolore come una modalità specifica separata di input sensoriale con il proprio apparato, mentre la `` teoria dei modelli '' riteneva che il dolore fosse il risultato dell'intensa stimolazione di recettori non specifici.2 Nel 1965, Wall e Melzack 3 La teoria del cancello del dolore ha fornito prove per un modello in cui la percezione del dolore era modulata sia dal feedback sensoriale che dal sistema nervoso centrale. Un altro enorme progresso nella teoria del dolore più o meno nello stesso periodo ha visto la scoperta della modalità di azione specifica degli oppioidi.4 Successivamente, i recenti progressi nella neuroimaging e nella medicina molecolare hanno notevolmente ampliato la nostra comprensione generale del dolore.
Quindi come si collega al dolore alla spalla? Il dolore alla spalla è un problema clinico comunee una solida comprensione del modo in cui il dolore viene elaborato dal corpo è essenziale per diagnosticare e trattare al meglio il dolore di un paziente. I progressi nella nostra conoscenza dell'elaborazione del dolore promettono di spiegare la mancata corrispondenza tra patologia e percezione del dolore, possono anche aiutarci a spiegare perché alcuni pazienti non rispondono a determinati trattamenti.
BLOCCHI DI COSTRUZIONE BASE DI DOLORE
Recettori sensoriali periferici: il meccanorecettore e l '' innocicettore '
Esistono numerosi tipi di recettori sensoriali periferici presenti nel sistema muscolo-scheletrico umano. 5 Possono essere classificati in base alla loro funzione (come meccanocettori, termocettori o nocicettori) o morfologia (terminazioni nervose libere o diversi tipi di recettori incapsulati) 5 I diversi tipi di recettori possono quindi essere ulteriormente sottoclassificati in base a la presenza di alcuni marcatori chimici. Ci sono sovrapposizioni significative tra diverse classi funzionali di recettori, per esempio
Elaborazione del dolore periferico: "Notociception"
Il danno tissutale coinvolge una varietà di mediatori infiammatori che vengono rilasciati da cellule danneggiate tra cui bradichinina, istamina, 5-idrossitriptamina, ATP, ossido nitrico e alcuni ioni (K + e H +). L'attivazione della via dell'acido arachidonico porta alla produzione di prostaglandine, trombossani e leucotrieni. Anche le citochine, comprese le interleuchine e il fattore di necrosi tumorale ?, e le neurotrofine, come il fattore di crescita nervoso (NGF), vengono rilasciate e sono intimamente coinvolte nella facilitazione dell'infiammazione.15 Altre sostanze come gli amminoacidi eccitatori (glutammato) e gli oppioidi ( endotelina-1) sono stati anche implicati nella risposta infiammatoria acuta.16 17 Alcuni di questi agenti possono attivare direttamente nocicettori, mentre altri determinano il reclutamento di altre cellule che poi rilasciano ulteriori agenti facilitatori.18 Questo processo locale che determina una maggiore reattività di neuroni nocicettivi al loro normale input e / o il reclutamento di una risposta a input normalmente sottosoglia è definito `` sensibilizzazione periferica ''. La Figura 1 riassume alcuni dei meccanismi chiave coinvolti.
NGF e il recettore transitorio del potenziale recettore del canale cationico sottofamiglia V membro 1 (TRPV1) hanno una relazione simbiotica quando si tratta di infiammazione e sensibilizzazione dei nocicettori. Le citochine prodotte nel tessuto infiammato determinano un aumento della produzione di NGF.19 L'NGF stimola il rilascio di istamina e serotonina (5-HT3) da parte dei mastociti e sensibilizza anche i nocicettori, alterando eventualmente le proprietà di A? fibre tali che una percentuale maggiore diventa nocicettiva. Il recettore TRPV1 è presente in una sottopopolazione di fibre afferenti primarie ed è attivato da capsaicina, calore e protoni. Il recettore TRPV1 è sintetizzato nel corpo cellulare della fibra afferente e viene trasportato sia al terminale periferico che a quello centrale, dove contribuisce alla sensibilità delle afferenze nocicettive. L'infiammazione provoca la produzione periferica di NGF che si lega quindi al recettore della tirosin chinasi di tipo 1 sui terminali dei nocicettori, l'NGF viene quindi trasportato al corpo cellulare dove porta a una regolazione della trascrizione di TRPV1 e di conseguenza aumenta la sensibilità dei nocicettori.19 20 NGF e altri mediatori infiammatori sensibilizzano anche TRPV1 attraverso una vasta gamma di vie di messaggistica secondarie. Si ritiene che molti altri recettori, inclusi i recettori colinergici, i recettori dell'acido α-aminobutirrico (GABA) e i recettori della somatostatina, siano coinvolti nella sensibilità dei nocicettori periferici.
Un gran numero di mediatori dell'infiammazione sono stati specificamente implicati nel dolore alla spalla e nella malattia della cuffia dei rotatori.21 Mentre alcuni mediatori chimici attivano direttamente i nocicettori, la maggior parte porta a cambiamenti nel neurone sensoriale stesso piuttosto che attivarlo direttamente. Questi cambiamenti possono dipendere dalla trascrizione post-traduzionale precoce o ritardata. Esempi del primo sono i cambiamenti nel recettore TRPV25 o nei canali ionici voltaggio-dipendenti risultanti dalla fosforilazione delle proteine legate alla membrana. Esempi di questi ultimi includono l'aumento indotto da NGF nella produzione del canale TRV1 e l'attivazione indotta dal calcio di fattori di trascrizione intracellulare.
Meccanismi molecolari di nocicezione
La sensazione di dolore ci avverte di lesioni reali o imminenti e innesca risposte protettive appropriate. Sfortunatamente, il dolore spesso sopravvive alla sua utilità come sistema di allarme e invece diventa cronico e debilitante. Questa transizione a una fase cronica comporta cambiamenti all'interno del midollo spinale e del cervello, ma c'è anche una notevole modulazione in cui i messaggi di dolore vengono avviati - a livello del neurone sensoriale primario. Gli sforzi per determinare come questi neuroni rilevano stimoli che producono dolore di natura termica, meccanica o chimica hanno rivelato nuovi meccanismi di segnalazione e ci hanno avvicinato alla comprensione degli eventi molecolari che facilitano le transizioni dal dolore acuto a quello persistente.
La neurochimica dei nocicettori
Il glutammato è il neurotrasmettitore eccitatorio predominante in tutti i nocicettori. Gli studi istochimici del DRG adulto, tuttavia, rivelano due ampie classi di fibra C non mielinizzata.
Trasduttori chimici per peggiorare il dolore
Come descritto sopra, il danno accresce la nostra esperienza dolorosa aumentando la sensibilità dei nocicettori agli stimoli sia termici che meccanici. Questo fenomeno deriva, in parte, dalla produzione e dal rilascio di mediatori chimici dal terminale sensoriale primario e da cellule non neurali (ad esempio, fibroblasti, mastociti, neutrofili e piastrine) nell'ambiente36 (Figura 3). Alcuni componenti del brodo infiammatorio (ad esempio protoni, ATP, serotonina o lipidi) possono alterare l'eccitabilità neuronale direttamente interagendo con i canali ionici sulla superficie del nocicettore, mentre altri (ad esempio, bradichinina e NGF) si legano ai recettori metabotropici e mediare i loro effetti attraverso le cascate di segnalazione secondo messenger11. Sono stati fatti notevoli progressi nella comprensione delle basi biochimiche di tali meccanismi modulatori.
Protoni extracellulari e acidosi dei tessuti
L'acidosi tessutale locale è una tipica risposta fisiologica alle lesioni e il grado di dolore o disagio associato è ben correlato con la grandezza dell'acidificazione37. L'applicazione di acido (pH 5) alla pelle produce scarichi sostenuti in un terzo o più dei nocicettori polimodali che innervano il campo recettivo 20.
Meccanismi cellulari e molecolari del dolore
Astratto
Il sistema nervoso rileva e interpreta una vasta gamma di stimoli termici e meccanici nonché sostanze irritanti chimiche ambientali ed endogene. Quando sono intensi, questi stimoli generano dolore acuto e, nel contesto di una lesione persistente, le componenti del sistema nervoso sia periferico che centrale del sistema di trasmissione del dolore mostrano una plasticità tremenda, aumentando i segnali del dolore e producendo ipersensibilità. Quando la plasticità facilita i riflessi protettivi, può essere utile, ma quando i cambiamenti persistono, può verificarsi una condizione di dolore cronico. Studi genetici, elettrofisiologici e farmacologici stanno chiarendo i meccanismi molecolari che sono alla base della rilevazione, della codifica e della modulazione di stimoli nocivi che generano dolore.
Introduzione: dolore acuto e persistente
Figura 5. Sensibilizzazione al midollo spinale (centrale)
Sensibilizzazione al recettore mediata da glutammato / NMDA.A seguito di stimolazione intensa o lesioni persistenti, C e A attivati? i nocicettori rilasciano una varietà di neurotrasmettitori tra cui dlutammato, sostanza P, peptide correlato al gene della calcitonina (CGRP) e ATP, sui neuroni di uscita nella lamina I del corno dorsale superficiale (rosso). Di conseguenza, i recettori del glutammato NMDA normalmente silenti situati nel neurone postsinaptico possono ora segnalare, aumentare il calcio intracellulare e attivare una serie di vie di segnalazione dipendenti dal calcio e secondi messaggeri tra cui la proteina chinasi attivata dal mitogeno (MAPK), la protein chinasi C (PKC) , proteina chinasi A (PKA) e Src. Questa cascata di eventi aumenterà l'eccitabilità del neurone di output e faciliterà la trasmissione dei messaggi di dolore al cervello.
Disinibizione.In circostanze normali, gli interneuroni inibitori (blu) rilasciano continuamente GABA e / o glicina (Gly) per diminuire l'eccitabilità dei neuroni di uscita della lamina I e modulare la trasmissione del dolore (tono inibitorio). Tuttavia, nel contesto della lesione, questa inibizione può essere persa, con conseguente iperalgesia. Inoltre, la disinibizione può abilitare mielinizzati A non nocicettivi? afferenze primarie per attivare i circuiti di trasmissione del dolore in modo tale che gli stimoli normalmente innocui vengono ora percepiti come dolorosi. Ciò avviene, in parte, attraverso la disinibizione della PKC eccitatoria? esprimendo interneuroni nella lamina interna II.
Attivazione microglialeLa lesione del nervo periferico promuove il rilascio di ATP e della chemochina fractalkine che stimolerà le cellule microgliali. In particolare, l'attivazione dei recettori purinergici, CX3CR1 e Toll-like sulla microglia (viola) si traduce nel rilascio del fattore neurotrofico derivato dal cervello (BDNF), che attraverso l'attivazione dei recettori TrkB espressi dai neuroni di uscita della lamina I, promuove una maggiore eccitabilità e aumento del dolore in risposta alla stimolazione sia nociva che innocua (cioè iperalgesia e allodinia). La microglia attivata rilascia anche una serie di citochine, come il fattore di necrosi tumorale? (TNF?), Interleuchina-1? e 6 (IL-1 ?, IL-6) e altri fattori che contribuiscono alla sensibilizzazione centrale.
Il milieu chimico di infiammazione
La sensibilizzazione periferica più comunemente deriva da cambiamenti associati all'infiammazione nell'ambiente chimico della fibra nervosa (McMahon et al., 2008). Pertanto, il danno tissutale è spesso accompagnato dall'accumulo di fattori endogeni rilasciati da nocicettori attivati o cellule non neurali che risiedono all'interno o si infiltrano nell'area danneggiata (inclusi mastociti, basofili, piastrine, macrofagi, neutrofili, cellule endoteliali, cheratinociti e fibroblasti). Collettivamente. questi fattori, denominati `` zuppa infiammatoria '', rappresentano una vasta gamma di molecole di segnalazione, inclusi neurotrasmettitori, peptidi (sostanza P, CGRP, bradichinina), eicosinoidi e lipidi correlati (prostaglandine, trombossani, leucotrieni, endocannabinoidi), neurotrofine, citochine e chemochine, così come proteasi e protoni extracellulari. Sorprendentemente, i nocicettori esprimono uno o più recettori della superficie cellulare in grado di riconoscere e rispondere a ciascuno di questi agenti pro-infiammatori o pro-algesici (Figura 4). Tali interazioni aumentano l'eccitabilità della fibra nervosa, aumentando così la sua sensibilità alla temperatura o al tatto.
Indiscutibilmente l'approccio più comune per ridurre il dolore infiammatorio comporta l'inibizione della sintesi o dell'accumulo di componenti del brodo infiammatorio. Questo è meglio esemplificato da farmaci antinfiammatori non steroidei, come l'aspirina o l'ibuprofene, che riducono il dolore infiammatorio e l'iperalgesia inibendo le cicloossigenasi (Cox-1 e Cox-2) coinvolte nella sintesi delle prostaglandine. Un secondo approccio consiste nel bloccare le azioni degli agenti infiammatori al nocicettore. Qui, evidenziamo esempi che forniscono nuove informazioni sui meccanismi cellulari di sensibilizzazione periferica o che costituiscono la base di nuove strategie terapeutiche per il trattamento del dolore infiammatorio.
L'NGF è forse meglio conosciuto per il suo ruolo come fattore neurotrofico necessario per la sopravvivenza e lo sviluppo dei neuroni sensoriali durante l'embriogenesi, ma nell'adulto, NGF è anche prodotto nel contesto di danno tissutale e costituisce una componente importante del brodo infiammatorio (Ritner et al., 2009). Tra i suoi numerosi bersagli cellulari, NGF agisce direttamente sui nocicettori della fibra C peptidergici, che esprimono la tirosin-chinasi del recettore NGF ad alta affinità, TrkA, così come il recettore della neurotropina a bassa affinità, p75 (Chao, 2003; Snider and McMahon, 1998). NGF produce una profonda ipersensibilità al calore e agli stimoli meccanici attraverso due meccanismi distinti temporalmente. Inizialmente, un'interazione NGF-TrkA attiva le vie di segnalazione a valle, tra cui la fosfolipasi C (PLC), la chinasi proteica attivata dal mitogeno (MAPK) e il fosfoinositide 3-chinasi (PI3K). Ciò si traduce in un potenziamento funzionale delle proteine bersaglio nel terminale periferico dei nocicettori, in particolare TRPV1, che porta a un rapido cambiamento della sensibilità al calore cellulare e comportamentale (Chuang et al., 2001).
Indipendentemente dai loro meccanismi pro-nocicettivi, l'interferenza con la segnalazione di neurotrofine o citochine è diventata una strategia importante per il controllo della malattia infiammatoria o del dolore risultante. L'approccio principale prevede il blocco di NGF o TNF-? azione con un anticorpo neutralizzante. Nel caso del TNF-?, Questo è stato notevolmente efficace nel trattamento di numerose malattie autoimmuni, inclusa l'artrite reumatoide, portando a una drastica riduzione sia della distruzione dei tessuti che dell'iperalgesia di accompagnamento (Atzeni et al., 2005). Poiché le principali azioni di NGF sul nocicettore adulto si verificano nel contesto dell'infiammazione, il vantaggio di questo approccio è che l'iperalgesia diminuirà senza influire normale percezione del dolore. Infatti, gli anticorpi anti-NGF sono attualmente in studi clinici per il trattamento delle sindromi da dolore infiammatorio (Hefti et al., 2006).
Sensibilizzazione mediata dal recettore glutammato / NMDA
Il dolore acuto è segnalato dal rilascio di glutammato dai terminali centrali dei nocicettori, generando correnti eccitatorie post-sinaptiche (EPSC) in neuroni del corno dorsale di secondo ordine. Ciò si verifica principalmente attraverso l'attivazione di AMPA postsinaptici e sottotipi di kainato di recettori del glutammato ionotropico. La sommatoria degli EPSC sub-soglia nel neurone postsinaptico finirà per provocare un potenziale di azione e la trasmissione del messaggio del dolore ai neuroni di ordine superiore.
Altri studi indicano che i cambiamenti nel neurone di proiezione, di per sé, contribuiscono al processo di inibizione. Ad esempio, la lesione del nervo periferico regola in modo profondo il trasportatore K + - Cl- KCC2, che è essenziale per mantenere i gradienti normali di K + e Cl attraverso la membrana plasmatica (Coull et al., 2003). La downregolazione del KCC2, che è espressa nei neuroni di proiezione della lamina I, determina uno spostamento del gradiente Cl, tale che l'attivazione dei recettori GABA-A depolarizza, piuttosto che iperpolarizzare i neuroni di proiezione della lamina I. Ciò, a sua volta, aumenterebbe l'eccitabilità e aumenterebbe la trasmissione del dolore. Infatti, il blocco farmacologico o la downregulation mediata da siRNA di KCC2 nel ratto induce allodinia meccanica.
Condividi Ebook
Fonte:
Perché mi fa male la spalla? Una revisione delle basi neuroanatomiche e biochimiche del dolore alla spalla
Benjamin John Floyd Dean, Stephen Edward Gwilym, Andrew Jonathan Carr
Meccanismi cellulari e molecolari del dolore
Allan I. Basbaum1, Diana M. Bautista2, Gre? Gory Scherrer1 e David Julius3
1Department of Anatomy, Università della California, San Francisco 94158
2Dipartimento di Biologia Molecolare e Cellulare, Università della California, Berkeley CA 94720 3Dipartimento di Fisiologia, Università della California, San Francisco 94158
Meccanismi molecolari della nocicezione
David Julius * e Allan I. Basbaum
*Dipartimento di Farmacologia Cellulare e Molecolare e �Dipartimenti di Anatomia e Fisiologia e WM Keck Foundation Center for Integrative Neuroscience, University of California San Francisco, San Francisco, California 94143, USA (e-mail: julius@socrates.ucsf.edu)
Infiammazione neurogena, o NI, è il processo fisiologico in cui i mediatori vengono scaricati direttamente dai nervi cutanei per iniziare una risposta infiammatoria. Ciò si traduce nella creazione di reazioni infiammatorie locali tra cui, eritema, gonfiore, aumento della temperatura, dolorabilità e dolore. Le sottili fibre somatiche afferenti non mielinizzate, che rispondono alle stimolazioni meccaniche e chimiche a bassa intensità, sono in gran parte responsabili del rilascio di questi mediatori dell'infiammazione.
Quando stimolati, queste vie nervose nei nervi cutanei rilasciano i neuropeptidi energetici, o sostanza P e il peptide correlato al gene della calcitonina (CGRP), rapidamente nel microambiente, innescando una serie di risposte infiammatorie. C'è una significativa distinzione nell'infiammazione immunogenica, che è la prima risposta protettiva e riparativa del sistema immunitario quando un agente patogeno entra nel corpo, mentre l'infiammazione neurogena implica una connessione diretta tra il sistema nervoso e le risposte infiammatorie. Anche se l'infiammazione neurogena e l'infiammazione immunologica possono esistere contemporaneamente, i due non sono clinicamente indistinguibili. Lo scopo di questo articolo è di discutere il meccanismo dell'infiammazione neurogena e il ruolo del sistema nervoso periferico nella difesa dell'ospite e nell'immunopatologia.
Infiammazione neurogena - Il ruolo del sistema nervoso periferico nella difesa dell'ospite e nell'immunopatologia
Astratto
Il sistema nervoso e il sistema immunitario periferico sono tradizionalmente considerati come funzioni separate. Questa linea, tuttavia, sta diventando sempre più offuscata dalle nuove conoscenze sull'infiammazione neurogena. I neuroni nocicettori possiedono molte delle stesse vie di riconoscimento molecolare per il pericolo delle cellule immunitarie e in risposta al pericolo, il sistema nervoso periferico comunica direttamente con il sistema immunitario, formando un meccanismo protettivo integrato. La fitta rete di innervazione delle fibre sensoriali e autonomiche nei tessuti periferici e l'alta velocità di trasduzione neurale consente una rapida modulazione neurogenica locale e sistemica dell'immunità. Anche i neuroni periferici sembrano giocare un ruolo significativo nella disfunzione immunitaria nelle malattie autoimmuni e allergiche. Pertanto, la comprensione dell'interazione coordinata dei neuroni periferici con le cellule immunitarie può far avanzare approcci terapeutici per aumentare la difesa dell'ospite e sopprimere l'immunopatologia.
Introduzione
Duemila anni fa, Celso definì l'infiammazione come coinvolgente quattro segni cardinali: Dolor (dolore), Calor (calore), Rubor (arrossamento) e Tumore (gonfiore), un'osservazione che indicava che l'attivazione del sistema nervoso era riconosciuta come parte integrante del infiammazione. Tuttavia, il dolore è stato pensato principalmente da allora, solo come un sintomo e non come un partecipante alla generazione dell'infiammazione. In questa prospettiva, dimostriamo che il sistema nervoso periferico svolge un ruolo diretto e attivo nel modulare l'immunità innata e adattativa, in modo tale che i sistemi immunitario e nervoso possano avere una funzione protettiva integrata comune nella difesa dell'ospite e nella risposta al danno tissutale, un intricato interazione che può anche portare a patologie nelle malattie allergiche e autoimmuni.
La sopravvivenza degli organismi dipende in modo critico dalla capacità di montare una difesa contro potenziali danni causati da danni ai tessuti e infezioni. La difesa dell'ospite coinvolge sia il comportamento di evitamento per rimuovere il contatto con un ambiente pericoloso (nocivo) (una funzione neurale), sia la neutralizzazione attiva dei patogeni (una funzione immunitaria). Tradizionalmente, il ruolo del sistema immunitario nella lotta agli agenti infettivi e nella riparazione delle lesioni tissutali è stato considerato abbastanza distinto da quello del sistema nervoso, che trasduce i segnali ambientali e interni dannosi in attività elettrica per produrre sensazioni e riflessi (Fig.1). Proponiamo che questi due sistemi siano effettivamente componenti di un meccanismo di difesa unificato. Il sistema nervoso somatosensoriale è nella posizione ideale per rilevare il pericolo. In primo luogo, tutti i tessuti altamente esposti all'ambiente esterno, come le superfici epiteliali della pelle, dei polmoni, del tratto urinario e digerente, sono densamente innervati dai nocicettori, fibre sensoriali ad alta soglia che producono dolore. In secondo luogo, la trasduzione di stimoli esterni nocivi è quasi istantanea, ordini di grandezza più rapida della mobilitazione del sistema immunitario innato, e quindi può essere il `` primo responder '' nella difesa dell'ospite.
Figura 1: Stimoli nocivi, percorsi di riconoscimento microbico e infiammatorio innescano l'attivazione del sistema nervoso periferico. I neuroni sensoriali possiedono diversi mezzi per rilevare la presenza di stimoli nocivi / nocivi. 1) I recettori del segnale di pericolo, compresi i canali TRP, i canali P2X ei recettori del pattern molecolare (DAMP) associati al pericolo riconoscono i segnali esogeni dall'ambiente (ad esempio calore, acidità, sostanze chimiche) o segnali di pericolo endogeni rilasciati durante il trauma / lesioni tissutali (ad es. ATP, acido urico, idrossinonali). 2) I recettori di riconoscimento del modello (PRR) come i recettori Toll-like (TLR) e recettori Nod-like (NLR) riconoscono i pattern molecolari associati al patogeno (PAMP) liberati dall'invasione di batteri o virus durante l'infezione. 3) I recettori delle citochine riconoscono i fattori secreti dalle cellule immunitarie (es. IL-1beta, TNF-alfa, NGF), che attivano le chinasi della mappa e altri meccanismi di segnalazione per aumentare l'eccitabilità della membrana.
Oltre agli input ortodromici al midollo spinale e al cervello dalla periferia, i potenziali d'azione nei neuroni nocicettori possono anche essere trasmessi antidromicamente ai punti di diramazione fino alla periferia, il riflesso dell'assone. Questi, insieme a depolarizzazioni locali sostenute, portano a un rilascio rapido e locale di mediatori neurali sia dagli assoni periferici che dai terminali (Fig. 2) 1. Esperimenti classici di Goltz (nel 1874) e di Bayliss (nel 1901) hanno mostrato che le radici dorsali elettricamente stimolanti induce vasodilatazione cutanea, che ha portato al concetto di infiammazione neurogena , indipendente da quella prodotta dal sistema immunitario (Fig.3).
Figura 2: I fattori neuronali rilasciati dai neuroni sensoriali nocicettori guidano direttamente la chemiotassi dei leucociti, l'emodinamica vascolare e la risposta immunitaria. Quando stimoli nocivi attivano segnali afferenti nei nervi sensoriali, vengono generati riflessi assonici antidromici che inducono il rilascio di neuropeptidi ai terminali periferici dei neuroni. Questi mediatori molecolari hanno diverse azioni infiammatorie: 1) chemiotassi e attivazione di neutrofili, macrofagi e linfociti nel sito di lesione e degranulazione di mastociti. 2) Segnalazione alle cellule endoteliali vascolari per aumentare il flusso sanguigno, perdite vascolari ed edema. Ciò consente anche un più facile reclutamento di leucociti infiammatori. 3) Adescamento delle cellule dendritiche per guidare la successiva differenziazione delle cellule T helper nei sottotipi Th2 o Th17.
Figura 3: Cronologia dei progressi nella comprensione degli aspetti neurogenici dell'infiammazione da Celso fino ai giorni nostri.
L'infiammazione neurogena è mediata dal rilascio dei neuropeptidi del peptide correlato al gene della calcitonina (CGRP) e della sostanza P (SP) dai nocicettori, che agiscono direttamente sulle cellule endoteliali vascolari e sulla muscolatura liscia 2�5. CGRP produce effetti di vasodilatazione 2, 3, mentre SP aumenta la permeabilità capillare portando a stravaso di plasma ed edema 4, 5, contribuendo al rubor, al calor e al tumore di Celso. Tuttavia, i nocicettori rilasciano molti neuropeptidi aggiuntivi (database online: www.neuropeptides.nl/), tra cui adrenomedullina, neurocinine A e B, peptide intestinale vasoattivo (VIP), neuropeptide (NPY) e peptide di rilascio della gastrina (GRP), nonché altri mediatori molecolari come glutammato, ossido nitrico (NO) e citochine come eotassina 6.
Ora ci rendiamo conto che i mediatori rilasciati dai neuroni sensoriali nella periferia non solo agiscono sul sistema vascolare, ma attraggono e attivano anche direttamente le cellule immunitarie innate (mastociti, cellule dendritiche) e le cellule immunitarie adattative (linfociti T) 7-12. Nel contesto acuto del danno tissutale, ipotizziamo che l'infiammazione neurogena sia protettiva, facilitando la guarigione fisiologica delle ferite e la difesa immunitaria contro i patogeni attivando e reclutando le cellule immunitarie. Tuttavia, tali comunicazioni neuroimmuni probabilmente svolgono anche un ruolo importante nella fisiopatologia delle malattie allergiche e autoimmuni amplificando le risposte immunitarie patologiche o disadattive. In modelli animali di artrite reumatoide, ad esempio, Levine e colleghi hanno dimostrato che la denervazione dell'articolazione porta a una notevole attenuazione dell'infiammazione, che dipende dall'espressione neurale della sostanza P 13, 14. In recenti studi sull'infiammazione delle vie aeree allergiche, colite e psoriasi, i neuroni sensoriali primari svolgono un ruolo centrale nell'avvio e nell'aumento dell'attivazione dell'immunità innata e adattativa 15-17.
Proponiamo quindi che il sistema nervoso periferico svolga non solo un ruolo passivo nella difesa dell'ospite (rilevamento di stimoli nocivi e avvio del comportamento di evitamento), ma anche un ruolo attivo in concerto con il sistema immunitario nel modulare le risposte e il stimoli, un ruolo che può essere sovvertito a contribuire alla malattia.
Percorsi di riconoscimento dei pericoli condivisi nei sistemi immunitari periferici nervosi e innati
I neuroni sensoriali periferici sono adattati a riconoscere il pericolo per l'organismo in virtù della loro sensibilità a stimoli chimici meccanici, termici e irritanti intensi (Fig. 1). I canali ionici del potenziale transitorio recettoriale (TRP) sono i mediatori molecolari più studiati della nocicezione, conducendo l'ingresso non selettivo dei cationi all'attivazione da parte di vari stimoli nocivi. TRPV1 viene attivato da alte temperature, pH basso e capsaicina, il componente irritante vallinoide del peperoncino 18. TRPA1 media l'individuazione di sostanze chimiche reattive tra cui irritanti ambientali come gas lacrimogeni e isotiocianati industriali 19, ma, cosa più importante, è anche attivato durante lesioni tissutali da segnali molecolari endogeni tra cui 4-idrossinonenale e prostaglandine 20, 21.
È interessante notare che i neuroni sensoriali condividono molte delle stesse vie del recettore del riconoscimento molecolare dei patogeni e dei pericoli come cellule immunitarie innate, che consentono loro anche di rilevare i patogeni (Fig. 1). Nel sistema immunitario, i patogeni microbici vengono rilevati dai recettori di riconoscimento del pattern (PRR) codificati germinali, che riconoscono pattern molecolari associati a patogeni esogeni ampiamente conservati (PAMP). I primi PRR da identificare erano membri della famiglia dei recettori toll-like (TLR), che si legano ai lieviti, ai componenti delle pareti cellulari derivate da batteri e all'RNA virale 22. Dopo l'attivazione della PRR, vengono attivati i percorsi di segnalazione a valle che inducono la produzione di citochine e l'attivazione dell'immunità adattativa. Oltre ai TLR, le cellule immunitarie innate vengono attivate durante lesioni tissutali da segnali di pericolo derivati endogeni, noti anche come pattern molecolari associati al danno (DAMP) o allarmi 23, 24. Questi segnali di pericolo includono HMGB1, acido urico e proteine da shock termico rilasciati dalle cellule morenti durante la necrosi, attivando le cellule immunitarie durante risposte infiammatorie non infettive.
I PRR inclusi i TLR 3, 4, 7 e 9 sono espressi dai neuroni nocicettori e la stimolazione da parte dei ligandi TLR porta all'induzione di correnti interne e alla sensibilizzazione dei nocicettori ad altri stimoli dolorosi 25. Inoltre, l'attivazione dei neuroni sensoriali da parte del ligando TLR27 imiquimod porta all'attivazione di una via sensoriale specifica per il prurito 7. Questi risultati indicano che il dolore e il prurito associati all'infezione possono essere in parte dovuti all'attivazione diretta dei neuroni da parte di fattori patogeni, che a loro volta attivare le cellule immunitarie attraverso il rilascio periferico di molecole di segnalazione neuronale.
Un importante DAMP / alarmin rilasciato durante il danno cellulare è l'ATP, che è riconosciuto dai recettori purinergici sia sui neuroni nocicettori che sulle cellule immunitarie 28-30. I recettori purinergici sono costituiti da due famiglie: recettori P2X, canali cationici legati al ligando e recettori P2Y, recettori accoppiati a proteine G. Nei neuroni nocicettori, il riconoscimento dell'ATP avviene attraverso P2X3, portando a correnti cationiche rapidamente densificanti e dolore 28, 30 (Fig.1), mentre i recettori P2Y contribuiscono all'attivazione dei nocicettori mediante la sensibilizzazione del TRP e dei canali del sodio voltaggio-dipendenti. Nei macrofagi, il legame dell'ATP ai recettori P2X7 porta all'iperpolarizzazione e all'attivazione a valle dell'inflammasoma, un complesso molecolare importante nella generazione di IL-1beta e IL-18 29. Pertanto, l'ATP è un potente segnale di pericolo che attiva sia i neuroni periferici che quelli innati. immunità durante la lesione, e alcune prove suggeriscono addirittura che i neuroni esprimano parti del macchinario molecolare dell'inflammasoma 31.
Il rovescio della medaglia dei segnali di pericolo nei nocicettori è il ruolo dei canali TRP nell'attivazione delle cellule immunitarie. TRPV2, un omologo di TRPV1 attivato da calore nocivo, è espresso ad alti livelli nelle cellule immunitarie innate 32. L'ablazione genetica di TRPV2 ha portato a difetti nella fagocitosi dei macrofagi e nella clearance delle infezioni batteriche 32. I mastociti esprimono anche i canali TRPV, che possono mediare direttamente la loro degranulazione 33. Resta da stabilire se i segnali di pericolo endogeno attivino le cellule immunitarie in modo simile ai nocicettori.
Un mezzo chiave di comunicazione tra cellule immunitarie e neuroni nocicettori è attraverso le citochine. Dopo l'attivazione dei recettori delle citochine, le vie di trasduzione del segnale vengono attivate nei neuroni sensoriali che portano alla fosforilazione a valle delle proteine di membrana tra cui TRP e canali voltaggio-dipendenti (Fig.1). La conseguente sensibilizzazione dei nocicettori significa che stimoli meccanici e termici normalmente innocui possono ora attivare i nocicettori. L'interleuchina 1 beta e il TNF-alfa sono due importanti citochine rilasciate dalle cellule immunitarie innate durante l'infiammazione. IL-1beta e TNF-alfa sono rilevati direttamente dai nocicettori che esprimono i recettori affini, inducono l'attivazione delle chinasi della mappa p38 che portano ad una maggiore eccitabilità di membrana 34. Anche il fattore di crescita nervoso (NGF) e la prostaglandina E (36) sono i principali mediatori dell'infiammazione rilasciati dalle cellule immunitarie che agiscono direttamente sui neuroni sensoriali periferici per provocare la sensibilizzazione. Un effetto importante della sensibilizzazione ai nocicettori da parte dei fattori immunitari è un aumento del rilascio di neuropeptidi ai terminali periferici che attivano ulteriormente le cellule immunitarie, inducendo così un ciclo di feedback positivo che guida e facilita l'infiammazione.
Controllo sensoriale del sistema nervoso di immunità innata e adattativa
Nelle prime fasi dell'infiammazione, i neuroni sensoriali segnalano ai mastociti e alle cellule dendritiche residenti nei tessuti, che sono cellule immunitarie innate importanti per avviare la risposta immunitaria (Fig. 2). Studi anatomici hanno mostrato un'apposizione diretta dei terminali con i mastociti, così come con le cellule dendritiche, e i neuropeptidi rilasciati dai nocicettori possono indurre degranulazione o produzione di citochine in queste cellule 7, 9, 37. Questa interazione gioca un ruolo importante nelle vie aeree allergiche infiammazioni e dermatiti 10 12.
Durante la fase effettrice dell'infiammazione, le cellule immunitarie devono trovare la loro strada verso il sito specifico della lesione. Molti mediatori rilasciati da neuroni sensoriali, neuropeptidi, chemochine e glutammato sono chemiotattici per neutrofili, eosinofili, macrofagi e cellule T e migliorano l'adesione endoteliale che facilita l'homing delle cellule immunitarie 6, 38-41 (Fig.2). Inoltre, alcune prove implicano che i neuroni possano partecipare direttamente alla fase effettrice, poiché i neuropeptidi stessi possono avere funzioni antimicrobiche dirette 42.
Le molecole di segnalazione di derivazione neuronale possono anche dirigere il tipo di infiammazione, contribuendo alla differenziazione o alla specificazione di diversi tipi di cellule T immunitarie adattive. Un antigene viene fagocitato ed elaborato da cellule immunitarie innate, che poi migrano al linfonodo più vicino e presentano il peptide antigenico ai linfociti T na ve. A seconda del tipo di antigene, delle molecole costimolatorie sulla cellula immunitaria innata e delle combinazioni di citochine specifiche, le cellule T na ve maturano in sottotipi specifici che meglio servono allo sforzo infiammatorio per eliminare lo stimolo patogeno. Le cellule T CD4, o cellule T helper (Th), possono essere suddivise in quattro gruppi principali, Th1, Th2, Th17 e cellule regolatrici T (Treg). Le cellule Th1 sono principalmente coinvolte nella regolazione delle risposte immunitarie ai microrganismi intracellulari e alle malattie autoimmuni organo-specifiche; I Th2 sono fondamentali per l'immunità contro i patogeni extracellulari, come gli elminti, e sono responsabili di malattie infiammatorie allergiche; Le cellule Th17 svolgono un ruolo centrale nella protezione contro le sfide microbiche, come batteri e funghi extracellulari; Le cellule Treg sono coinvolte nel mantenimento dell'autotolleranza e nella regolazione delle risposte immunitarie. Questo processo di maturazione delle cellule T sembra essere fortemente influenzato dai mediatori neuronali sensoriali. I neuropeptidi, come CGRP e VIP, possono polarizzare le cellule dendritiche verso un'immunità di tipo Th2 e ridurre l'immunità di tipo Th1 promuovendo la produzione di alcune citochine e inibendone altre, nonché riducendo o migliorando la migrazione delle cellule dendritiche ai linfonodi locali 8 , 10, 43. I neuroni sensoriali contribuiscono anche considerevolmente all'infiammazione allergica (principalmente guidata da Th2) 17. Oltre a regolare le cellule Th1 e Th2, altri neuropeptidi, come SP e Hemokinin-1, possono guidare la risposta infiammatoria più verso Th17 o Treg 44, 45, il che significa che i neuroni possono anche essere coinvolti nella regolazione della risoluzione infiammatoria. Nelle immunopatologie come la colite e la psoriasi, il blocco dei mediatori neuronali come la sostanza P può smorzare in modo significativo i linfociti T e il danno immunomediato 15-17, sebbene antagonizzare un mediatore possa di per sé avere solo un effetto limitato sull'infiammazione neurogena.
Considerando che le molecole di segnalazione rilasciate dalle fibre nervose sensoriali periferiche regolano non solo i piccoli vasi sanguigni, ma anche la chemiotassi, l'homing, la maturazione e l'attivazione delle cellule immunitarie, sta diventando chiaro che le interazioni neuro-immuni sono molto più complesse di quanto si pensasse (Fig. 2). Inoltre, è abbastanza plausibile che non siano i singoli mediatori neurali, ma piuttosto combinazioni specifiche di molecole di segnalazione rilasciate dai nocicettori che influenzano diversi stadi e tipi di risposte immunitarie.
Controllo del riflesso autonomo dell'immunità
Appare anche prominente un ruolo per un circuito di "riflesso" del sistema nervoso autonomo colinergico nella regolazione delle risposte immunitarie periferiche 46. Il vago è il principale nervo parasimpatico che collega il tronco cerebrale con gli organi viscerali. Il lavoro di Kevin Tracey e altri indicano potenti risposte antinfiammatorie generalizzate nello shock settico e nell'endotossiemia, innescate da un'attività efferente del nervo vagale che porta a una soppressione dei macrofagi periferici 47. Il vago attiva i neuroni gangliari celiaci periferici che innervano la milza, portando al rilascio a valle di acetilcolina, che si lega ai recettori nicotinici alfa-49 sui macrofagi nella milza e nel tratto gastrointestinale. Ciò induce l'attivazione della via di segnalazione SOCS7 JAK2 / STAT3, che sopprime potentemente la trascrizione del TNF-alfa 3. Il ganglio adrenergico celiaco comunica anche direttamente con un sottoinsieme di cellule T della memoria che producono acetilcolina, che sopprimono i macrofagi infiammatori 47.
Le cellule T killer naturali invarianti (iNKT) sono un sottoinsieme specializzato di cellule T che riconoscono i lipidi microbici nel contesto di CD1d invece di antigeni peptidici. Le cellule NKT sono una popolazione chiave di linfociti coinvolti nel combattimento di agenti patogeni infettivi e nella regolazione dell'immunità sistemica. Le cellule NKT risiedono e si spostano principalmente attraverso la vascolatura e le sinusoidi della milza e del fegato. I nervi simpatici beta-adrenergici nel fegato segnalano direttamente per modulare l'attività delle cellule NKT 50. Nel corso di un modello murino di ictus (MCAO), ad esempio, la mobilità delle cellule NKT del fegato è stata visibilmente soppressa, che è stata invertita da denervazione simpatica o antagonisti beta-adrenergici. Inoltre, questa attività immunosoppressiva dei neuroni noradrenergici sulle cellule NKT ha portato ad un aumento delle infezioni sistemiche e delle lesioni polmonari. Pertanto, i segnali efferenti dei neuroni autonomi possono mediare una potente immuno-soppressione.
Insight di Dr. Alex Jimenez
L'infiammazione neurogena è una risposta infiammatoria locale generata dal sistema nervoso. Si ritiene che svolga un ruolo fondamentale nella patogenesi di una varietà di problemi di salute, tra cui emicrania, psoriasi, asma, fibromialgia, eczema, rosacea, distonia e sensibilità chimica multipla. Sebbene l'infiammazione neurogena associata al sistema nervoso periferico sia stata ampiamente studiata, il concetto di infiammazione neurogena all'interno del sistema nervoso centrale necessita ancora di ulteriori ricerche. Secondo diversi studi di ricerca, tuttavia, si ritiene che le carenze di magnesio siano la causa principale dell'infiammazione neurogena. Il seguente articolo mostra una panoramica dei meccanismi dell'infiammazione neurogena nel sistema nervoso, che può aiutare gli operatori sanitari a determinare il miglior approccio terapeutico per la cura di una varietà di problemi di salute associati al sistema nervoso.
Conclusioni
Quali sono i rispettivi ruoli specifici dei sistemi nervoso somatosensoriale e autonomo nella regolazione dell'infiammazione e del sistema immunitario (Fig.4)? L'attivazione dei nocicettori porta ai riflessi assonici locali, che reclutano e attivano localmente le cellule immunitarie ed è quindi principalmente pro-infiammatori e spazialmente confinati. Al contrario, la stimolazione autonomica porta a un'immunosoppressione sistemica influenzando i pool di cellule immunitarie nel fegato e nella milza. I meccanismi di segnalazione afferente nella periferia che portano all'attivazione del circuito riflesso colinergico vagale immunosoppressore sono poco conosciuti. Tuttavia, l'80-90% delle fibre vagali sono fibre sensoriali afferenti primarie, e quindi i segnali provenienti dai visceri, molti potenzialmente guidati dalle cellule immunitarie, possono portare all'attivazione degli interneuroni nel tronco cerebrale e attraverso di essi a un'uscita nelle fibre vagali efferenti 46.
Figura 4: I sistemi nervosi sensoriali e autonomi modulano rispettivamente le risposte immunitarie locali e sistemiche. I nocicettori che innervano le superfici epiteliali (ad es. Pelle e polmone) inducono risposte infiammatorie localizzate, attivando mastociti e cellule dendritiche. Nell'infiammazione delle vie aeree allergiche, nella dermatite e nell'artrite reumatoide, i neuroni nocicettori svolgono un ruolo nel guidare l'infiammazione. Al contrario, i circuiti autonomici che innervano gli organi viscerali (ad esempio milza e fegato) regolano le risposte immunitarie sistemiche bloccando l'attivazione delle cellule NKT e dei macrofagi. In endotossemia cerebrale e settica, questi neuroni svolgono un ruolo immunosoppressivo.
In genere, il corso del tempo e la natura dell'infiammazione, sia durante l'infezione, le reazioni allergiche o patologie autoimmuni, sono definiti dalle categorie di cellule immunitarie coinvolte. Sarà importante sapere quali diversi tipi di cellule immunitarie sono regolate da segnali sensoriali e autonomi. Una valutazione sistematica di ciò che i mediatori possono essere rilasciati dai nocicettori e dai neuroni autonomici e l'espressione dei recettori per questi da differenti cellule immunitarie innate e adattative potrebbe aiutare a rispondere a questa domanda.
Durante l'evoluzione, si sono sviluppati percorsi molecolari di rilevamento di pericolo simili sia per l'immunità innata che per la nocicezione anche se le cellule hanno linee di sviluppo completamente diverse. Mentre i PRR e i canali ionici legati ai leganti nocivi sono studiati separatamente da immunologi e neurobiologi, il confine tra questi due campi è sempre più sfocato. Durante il danno tissutale e l'infezione patogena, è probabile che il rilascio di segnali di pericolo porti ad un'attivazione coordinata sia dei neuroni periferici che delle cellule immunitarie con una comunicazione bidirezionale complessa e una difesa dell'ospite integrata. Il posizionamento anatomico dei nocicettori all'interfaccia con l'ambiente, la velocità di trasduzione neurale e la loro capacità di rilasciare potenti cocktail di mediatori ad azione immunitaria consentono al sistema nervoso periferico di modulare attivamente la risposta immunitaria innata e coordinare l'immunità adattativa a valle. Al contrario, i nocicettori sono altamente sensibili ai mediatori immunitari, che attivano e sensibilizzano i neuroni. L'infiammazione neurogena e quella immunomediata non sono, quindi, entità indipendenti, ma agiscono insieme come dispositivi di allarme precoce. Tuttavia, il sistema nervoso periferico svolge anche un ruolo importante nella fisiopatologia, e forse eziologia, di molte malattie immunitarie come asma, psoriasi o colite perché la sua capacità di attivare il sistema immunitario può amplificare l'infiammazione patologica 15-17. Il trattamento per i disturbi immunitari può richiedere, quindi, di includere il targeting dei nocicettori e delle cellule immunitarie.
Ringraziamenti
Ringraziamo il NIH per il supporto (2R37NS039518).
In conclusione,Comprendere il ruolo dell'infiammazione neurogena quando si tratta di difesa dell'ospite e immunopatologia è essenziale per determinare l'approccio terapeutico appropriato per una varietà di problemi di salute del sistema nervoso. Osservando le interazioni dei neuroni periferici con le cellule immunitarie, gli operatori sanitari possono far progredire approcci terapeutici per aumentare ulteriormente la difesa dell'ospite e sopprimere l'immunopatologia. Lo scopo dell'articolo sopra è quello di aiutare i pazienti a comprendere la neurofisiologia clinica della neuropatia, tra gli altri problemi di salute delle lesioni nervose. Informazioni referenziate dal National Center for Biotechnology Information (NCBI). Lo scopo delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
1. Sauer SK, Reeh PW, Bove GM. Rilascio di CGRP nocivo indotto dal calore dagli assoni del nervo sciatico di ratto in vitroEur J Neurosci. 2001;14: 1203 1208. [PubMed]
2. Edvinsson L, Ekman R, Jansen I, McCulloch J, Uddman R. Peptide correlato al gene della calcitonina e vasi sanguigni cerebrali: distribuzione ed effetti vasomotori.J Cereb Blood Flow Metab. 1987;7: 720 728. [PubMed]
3. McCormack DG, Mak JC, Coupe MO, Barnes PJ. Vasodilatazione peptidica correlata al gene della calcitonina dei vasi polmonari umaniJ Appl Physiol. 1989;67: 1265 1270. [PubMed]
4. Saria A. La sostanza P nelle fibre nervose sensoriali contribuisce allo sviluppo di edema nella zampa posteriore del ratto dopo una lesione termica.Br J Pharmacol. 1984;82: 217 222. [Articolo gratuito di PMC]ï ¿½[PubMed]
5. Cervello SD, Williams TJ. Le interazioni tra le tachichinine e il peptide generato dalla calcitonina portano alla modulazione della formazione di edema e del flusso sanguigno nella pelle di ratto.Br J Pharmacol. 1989;97: 77 82.[Articolo gratuito di PMC]ï ¿½[PubMed]
6. Fryer AD, et al. L'eotassina neuronale e gli effetti dell'antagonista CCR3 sull'iperreattività delle vie aeree e sulla disfunzione del recettore M2.J Clin Invest. 2006;116: 228 236. [Articolo gratuito di PMC]ï ¿½[PubMed]
7. Ansel JC, Brown JR, Payan DG, Brown MA. La sostanza P attiva selettivamente l'espressione genica del TNF-alfa nei mastociti muriniJ Immunol. 1993;150: 4478 4485. [PubMed]
8. Ding W, Stohl LL, Wagner JA, Granstein RD. Il peptide correlato al gene della calcitonina polarizza le cellule di Langerhans verso l'immunità di tipo Th2J Immunol. 2008;181: 6020 6026. [Articolo gratuito di PMC]ï ¿½[PubMed]
9. Hosoi J, et al. Regolazione della funzione delle cellule di Langerhans da parte dei nervi contenenti peptide correlato al gene della calcitoninaNatura1993;363: 159 163. [PubMed]
10. Mikami N, et al. Il peptide correlato al gene della calcitonina è un importante regolatore dell'immunità cutanea: effetto sulle funzioni delle cellule dendritiche e delle cellule T.J Immunol. 2011;186: 6886 6893. [PubMed]
11. Rochlitzer S, et al. Il peptide correlato al gene della calcitonina neuropeptide influenza l'infiammazione delle vie aeree allergiche modulando la funzione delle cellule dendritiche.Clin Exp Allergy2011;41: 1609 1621. [PubMed]
12. Cyphert JM, et al. La cooperazione tra mastociti e neuroni è essenziale per la broncocostrizione mediata dall'antigeneJ Immunol. 2009;182: 7430 7439. [Articolo gratuito di PMC]ï ¿½[PubMed]
13. Levine JD, et al. La sostanza intraneuronale P contribuisce alla gravità dell'artrite sperimentaleScienza1984;226: 547 549. [PubMed]
14. Levine JD, Khasar SG, Green PG. Infiammazione neurogena e artriteAnn NY Acad Sci. 2006;1069: 155 167. [PubMed]
15. Engel MA, et al. TRPA1 e la sostanza P mediano la colite nei topiGastroenterologia . 2011;141: 1346 1358. [PubMed]
16. Ostrowski SM, Belkadi A, Loyd CM, Diaconu D, Ward NL. La denervazione cutanea della pelle di topo psoriasiforme migliora l'acantosi e l'infiammazione in modo dipendente dal neuropeptide sensoriale.J Invest Dermatol. 2011;131: 1530 1538. [Articolo gratuito di PMC]ï ¿½[PubMed]
17. Caceres AI, et al. Un canale ionico neuronale sensoriale essenziale per l'infiammazione delle vie aeree e l'iperreattività nell'asmaProc Natl Acad Sci US A. 2009;106: 9099 9104. [Articolo gratuito di PMC]ï ¿½[PubMed]
18. Caterina MJ, et al. Nocicezione compromessa e sensazione di dolore nei topi privi del recettore della capsaicinaScienza2000;288: 306 313. [PubMed]
19. Bessac BF, et al. Gli antagonisti transitori del potenziale recettore ankyrin 1 bloccano gli effetti nocivi degli isocianati industriali tossici e dei gas lacrimogeni.FASEB J. 2009;23: 1102 1114. [Articolo gratuito di PMC]ï ¿½[PubMed]
20. Cruz-Orengo L, et al. Nocicezione cutanea evocata da 15-delta PGJ2 tramite l'attivazione del canale ionico TRPA1Mol Pain2008;4: 30. [Articolo gratuito di PMC]ï ¿½[PubMed]
21. Trevisani M, et al. 4-Hydroxynonenal, un'aldeide endogena, provoca dolore e infiammazione neurogena attraverso l'attivazione del recettore irritante TRPA1.Proc Natl Acad Sci US A. 2007;104: 13519 13524. [Articolo gratuito di PMC]ï ¿½[PubMed]
22. Janeway CA, Jr, Medzhitov R. Introduzione: il ruolo dell'immunità innata nella risposta immunitaria adattativa.Semin Immunol. 1998;10: 349 350. [PubMed]
23. Matzinger P.Un innato senso di pericoloAnn NY Acad Sci. 2002;961: 341 342. [PubMed]
24. Bianchi ME. DAMP, PAMP e allarmi: tutto ciò che dobbiamo sapere sul pericoloJ Leukoc Biol. 2007;81: 1 5. [PubMed]
25. Liu T, Xu ZZ, Park CK, Berta T, Ji RR. Il recettore simile al pedaggio 7 media il pruritoNat Neurosci. 2010;13: 1460 1462. [Articolo gratuito di PMC]ï ¿½[PubMed]
26. Diogene A, Ferraz CC, Akopian AN, Henry MA, Hargreaves KM. LPS sensibilizza TRPV1 tramite l'attivazione di TLR4 nei neuroni sensoriali trigeminaliJ Dent Res. 2011;90: 759 764. [PubMed]
27. Qi J, et al. Vie dolorose indotte dalla stimolazione TLR dei neuroni gangliari della radice dorsaleJ Immunol. 2011;186: 6417 6426. [Articolo gratuito di PMC]ï ¿½[PubMed]
28. Cockayne DA, et al. Iporeflessia della vescica urinaria e ridotto comportamento correlato al dolore nei topi con deficit di P2X3Natura2000;407: 1011 1015. [PubMed]
29. Mariathasan S, et al. La criopirina attiva l'inflammasoma in risposta alle tossine e all'ATPNatura2006;440: 228 232. [PubMed]
30. Souslova V, et al. Deficit di codifica a caldo e dolore infiammatorio aberrante nei topi privi di recettori P2X3Natura2000;407: 1015 1017. [PubMed]
31. de Rivero Vaccari JP, Lotocki G, Marcillo AE, Dietrich WD, Keane RW. Una piattaforma molecolare nei neuroni regola l'infiammazione dopo la lesione del midollo spinaleJ Neurosci. 2008;28: 3404 3414. [PubMed]
32. Link TM, et al. TRPV2 ha un ruolo fondamentale nel legame delle particelle dei macrofagi e nella fagocitosiNat Immunol. 2010;11: 232 239. [Articolo gratuito di PMC]ï ¿½[PubMed]
33. Turner H, del Carmen KA, Stokes A. Collegamento tra i canali TRPV e la funzione dei mastocitiHandb Exp Pharmacol. 2007: 457 471. [PubMed]
34. Binshtok AM, et al. I nocicettori sono sensori dell'interleuchina-1betaJ Neurosci. 2008;28: 14062 14073.[Articolo gratuito di PMC]ï ¿½[PubMed]
35. Zhang XC, Kainz V, Burstein R, Levy D.Il fattore alfa di necrosi tumorale induce la sensibilizzazione dei nocicettori meningei mediati tramite azioni COX locali e p38 MAP chinasi.Dolore2011;152: 140 149.[Articolo gratuito di PMC]ï ¿½[PubMed]
36. Samad TA, et al. L'induzione mediata dall'interleuchina-1beta di Cox-2 nel SNC contribuisce all'ipersensibilità al dolore infiammatorio.Natura2001;410: 471 475. [PubMed]
37. Veres TZ, et al. Interazioni spaziali tra cellule dendritiche e nervi sensoriali nell'infiammazione allergica delle vie aereeSono J Respir Cell Mol Biol. 2007;37: 553 561. [PubMed]
38. Smith CH, Barker JN, Morris RW, MacDonald DM, Lee TH. I neuropeptidi inducono una rapida espressione delle molecole di adesione delle cellule endoteliali e provocano l'infiltrazione granulocitica nella pelle umana.J Immunol. 1993;151: 3274 3282. [PubMed]
39. Dunzendorfer S, Meierhofer C, Wiedermann CJ. Segnalazione nella migrazione indotta da neuropeptidi di eosinofili umaniJ Leukoc Biol. 1998;64: 828 834. [PubMed]
40. Ganor Y, Besser M, Ben-Zakay N, Unger T, Levite M. Le cellule T umane esprimono un recettore ionotropico funzionale del glutammato GluR3 e il glutammato da solo innesca l'adesione mediata dall'integrina alla laminina e alla fibronectina e la migrazione chemiotattica.J Immunol. 2003;170: 4362 4372. [PubMed]
41. Czepielewski RS, et al. Il recettore del peptide di rilascio della gastrina (GRPR) media la chemiotassi nei neutrofiliProc Natl Acad Sci US A. 2011;109: 547 552. [Articolo gratuito di PMC]ï ¿½[PubMed]
42. Brogden KA, Guthmiller JM, Salzet M, Zasloff M. Il sistema nervoso e l'immunità innata: la connessione neuropeptidica.Nat Immunol. 2005;6: 558 564. [PubMed]
43. Jimeno R, et al. Effetto del VIP sull'equilibrio tra citochine e regolatori principali dei linfociti T helper attivatiImmunol Cell Biol. 2011;90: 178 186. [PubMed]
44. Razavi R, et al. I neuroni sensoriali TRPV1 + controllano lo stress delle cellule beta e l'infiammazione delle isole nel diabete autoimmuneCell. 2006;127: 1123 1135. [PubMed]
45. Cunin P, et al. La sostanza delle tachichinine P e l'emocinina-1 favoriscono la generazione di cellule Th17 della memoria umana inducendo l'espressione di IL-1beta, IL-23 e 1A simile al TNF da parte dei monociti.J Immunol. 2011;186: 4175 4182. [PubMed]
46. Andersson U, Tracey KJ. Principi riflessi dell'omeostasi immunologicaAnnu Rev Immunol. 2011[Articolo gratuito di PMC]ï ¿½[PubMed]
47. de Jonge WJ, et al. La stimolazione del nervo vago attenua l'attivazione dei macrofagi attivando la via di segnalazione Jak2-STAT3.Nat Immunol. 2005;6: 844 851. [PubMed]
48. Rosas-Ballina M, et al. Le cellule T che sintetizzano l'acetilcolina trasmettono segnali neurali in un circuito del nervo vagoScienza2011;334: 98 101. [Articolo gratuito di PMC]ï ¿½[PubMed]
49. Wang H, et al. La subunità alfa7 del recettore nicotinico dell'acetilcolina è un regolatore essenziale dell'infiammazioneNatura2003;421: 384 388. [PubMed]
50. Wong CH, Jenne CN, Lee WY, Leger C, Kubes P. L'innervazione funzionale delle cellule iNKT epatiche è immunosoppressiva dopo l'ictus.Scienza2011;334: 101 105. [PubMed]
Lo strumento Find A Practitioner di IFM è la più grande rete di riferimento in Medicina Funzionale, creata per aiutare i pazienti a localizzare professionisti di Medicina Funzionale in qualsiasi parte del mondo. I professionisti certificati IFM sono elencati per primi nei risultati di ricerca, data la loro vasta formazione in Medicina Funzionale
Prenotazione online e appuntamenti 24 ore su 7, XNUMX giorni su XNUMX *



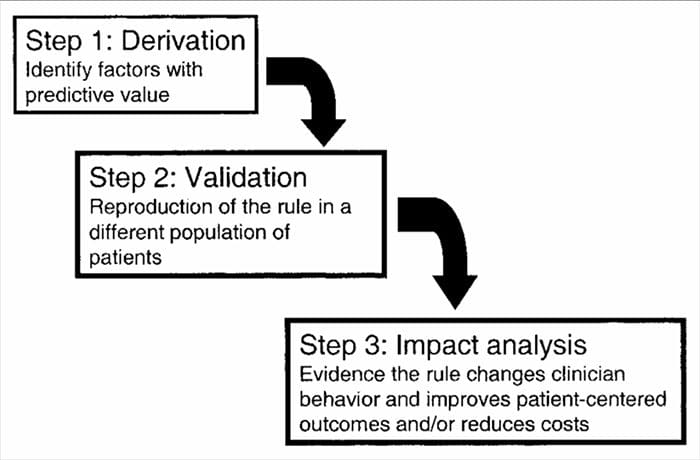




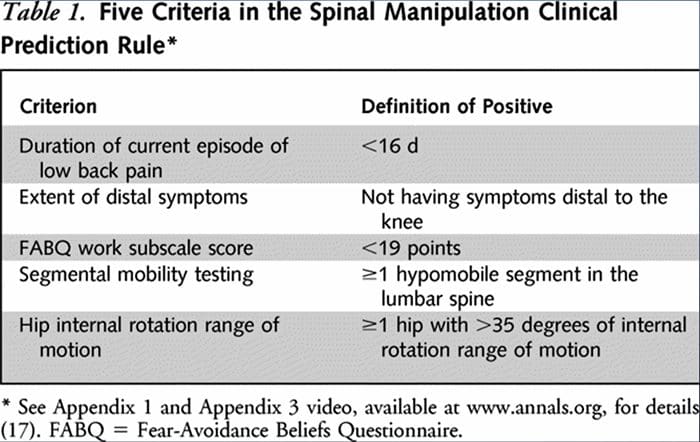
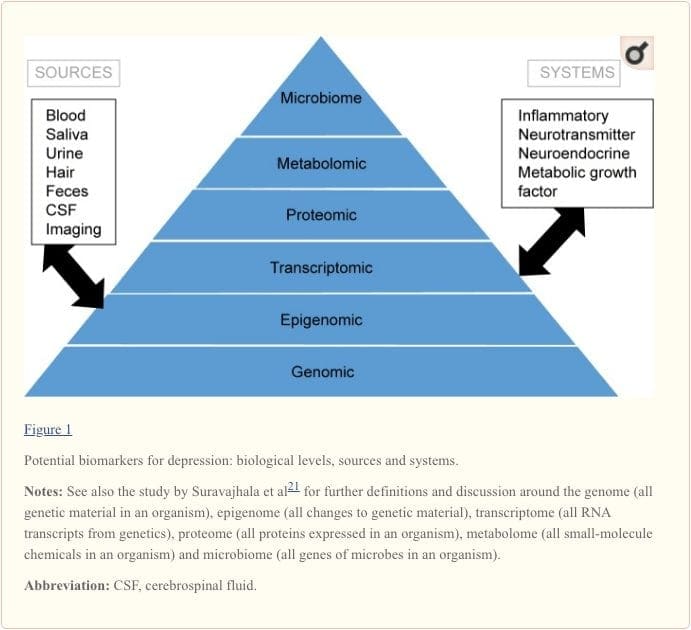





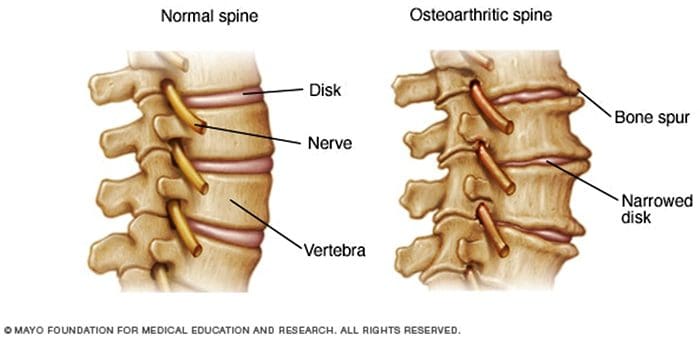
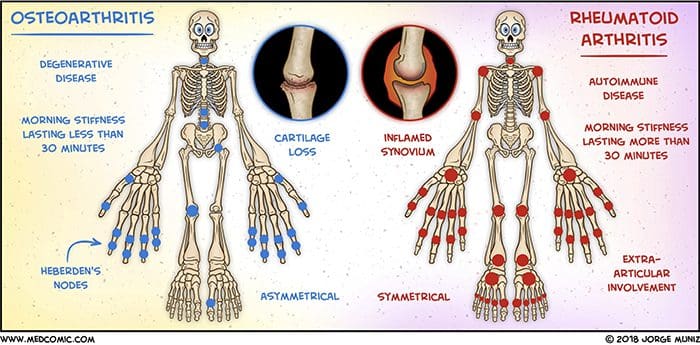 Astratto
Astratto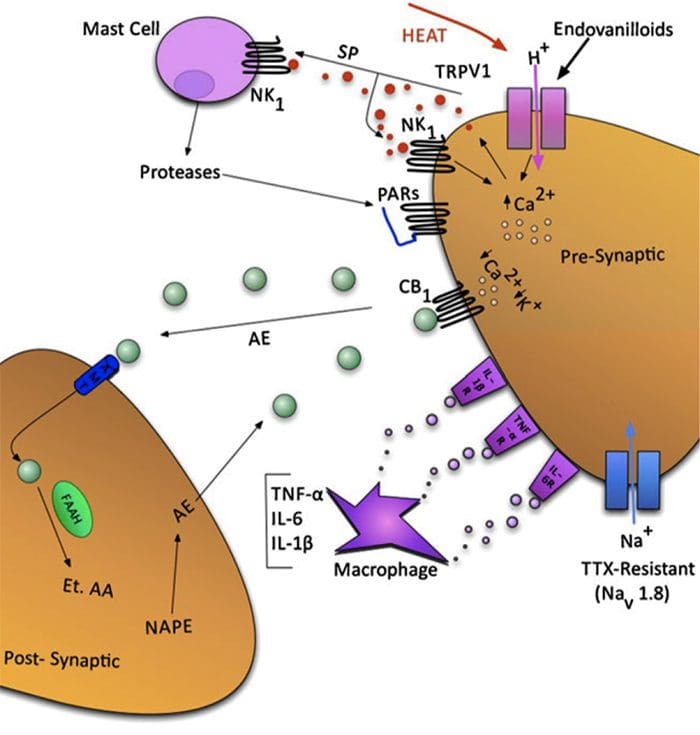
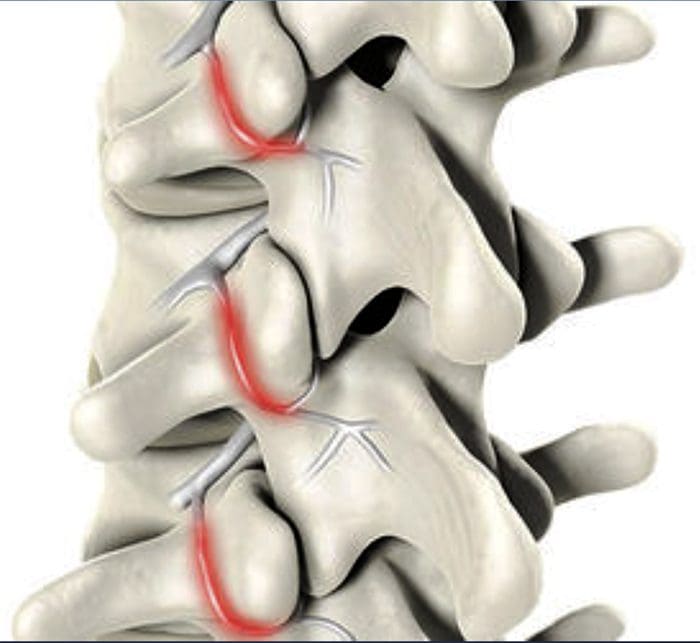
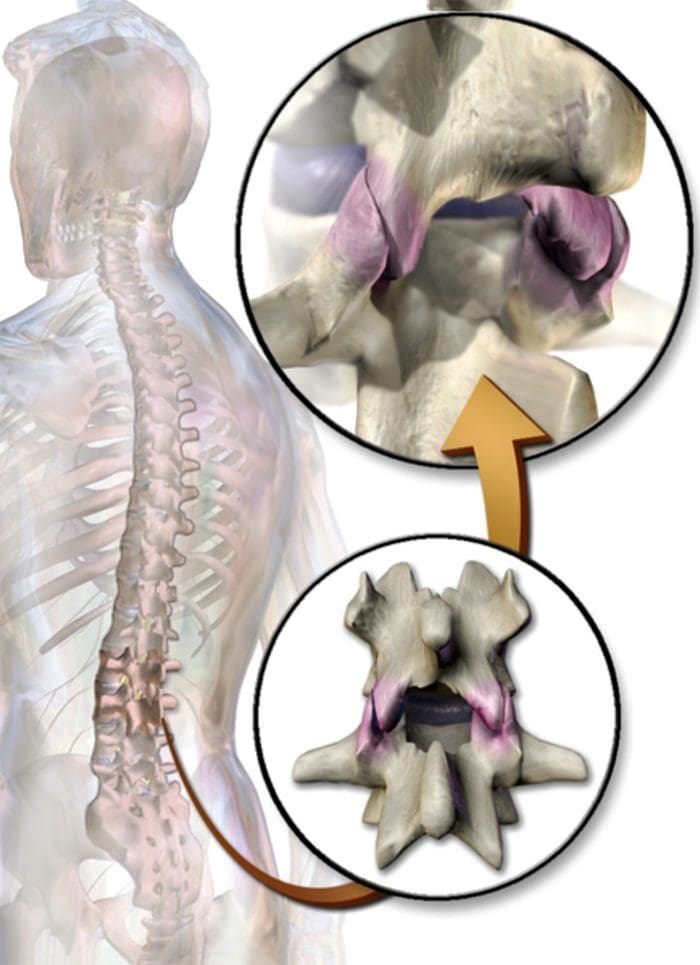
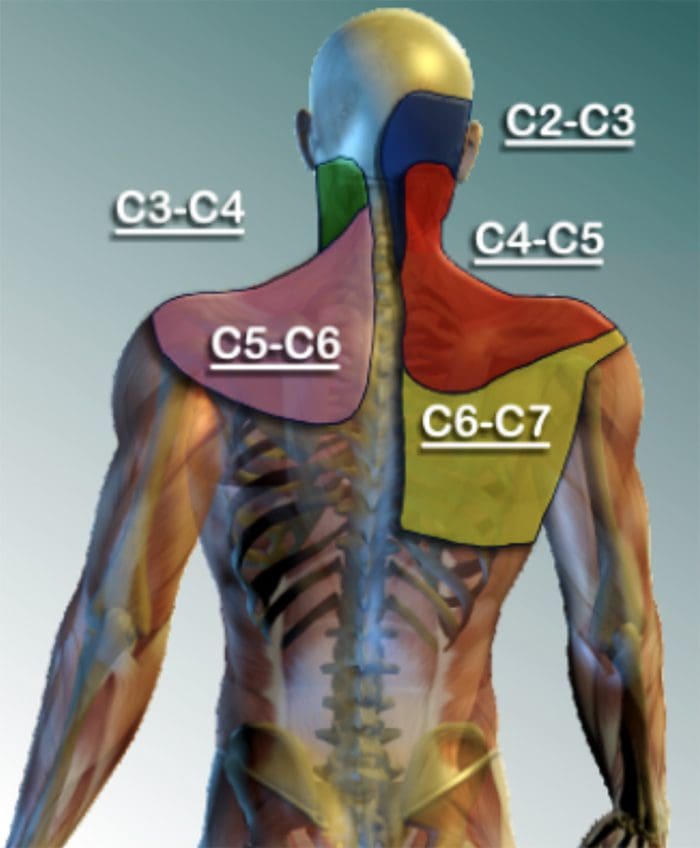
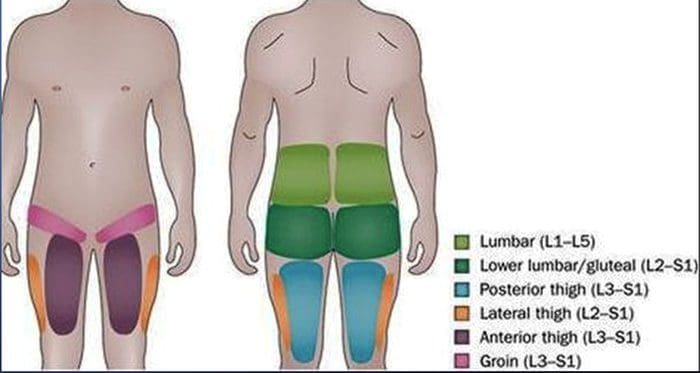

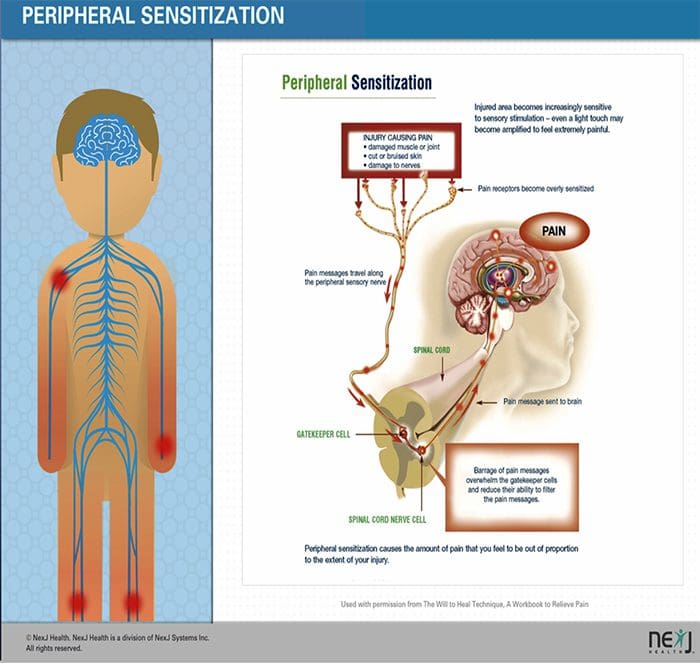
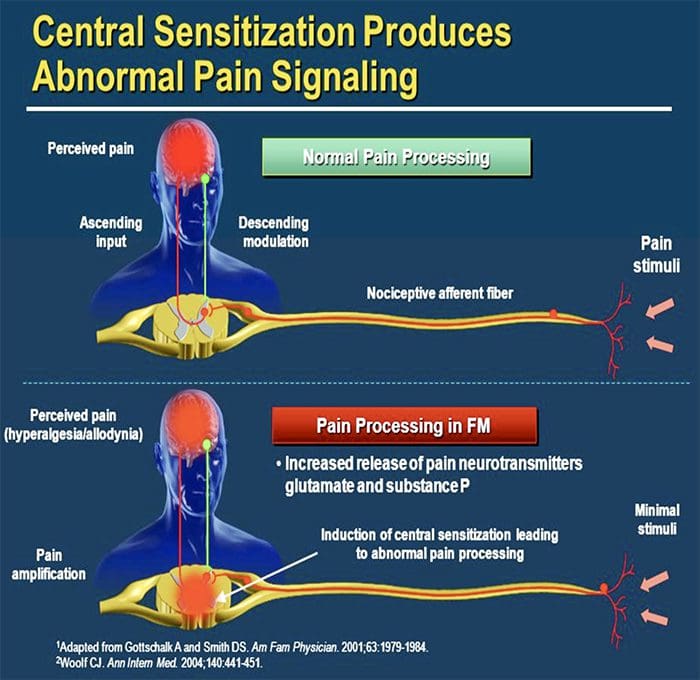


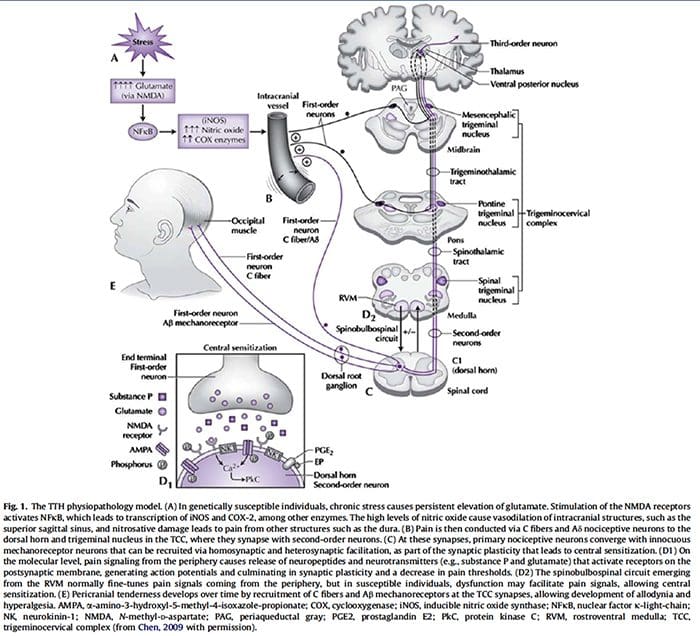

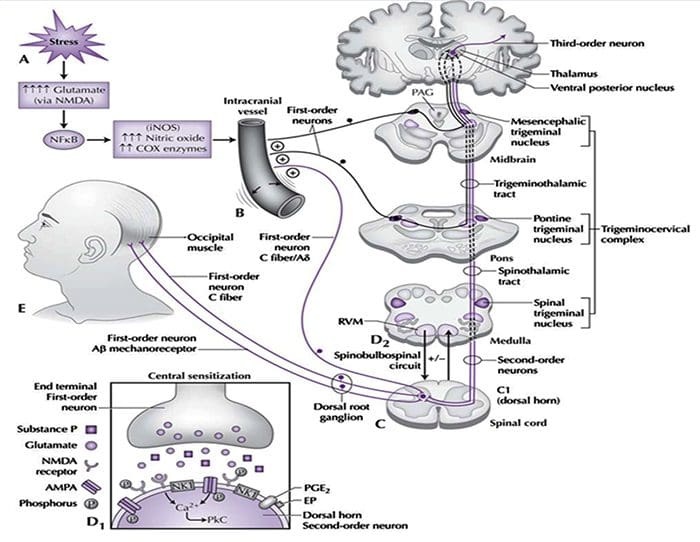




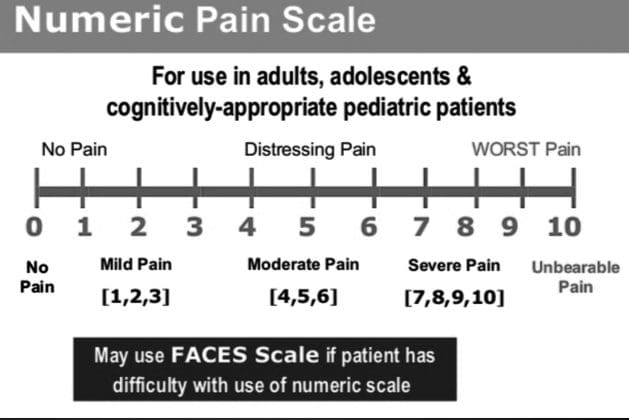








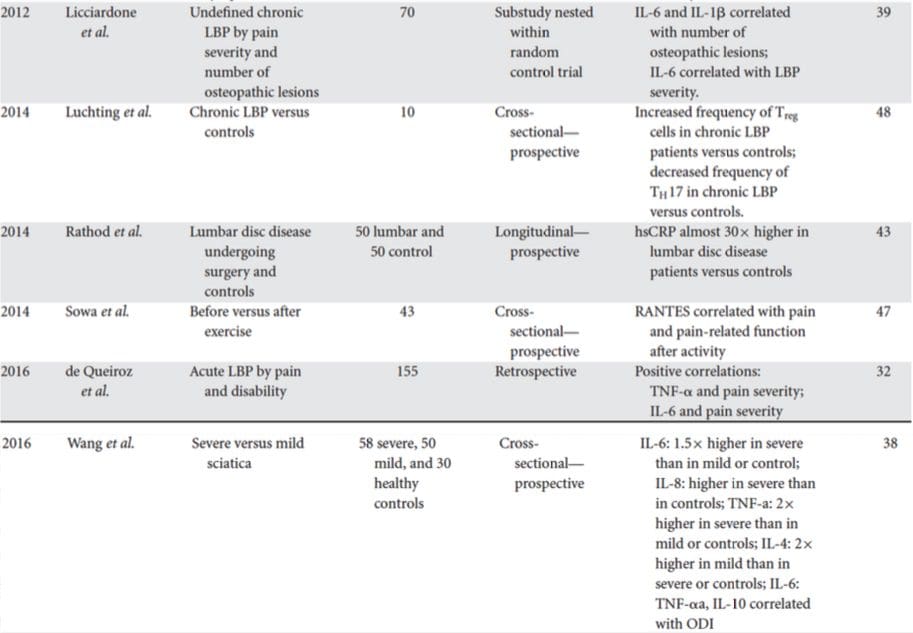


 Riferimenti:
Riferimenti:
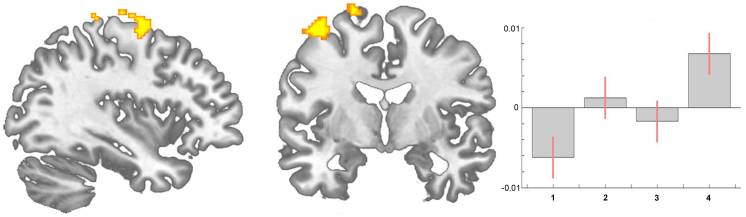
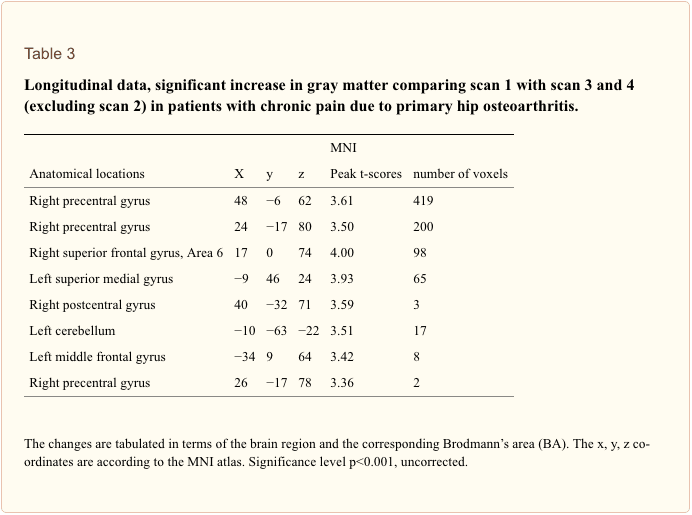

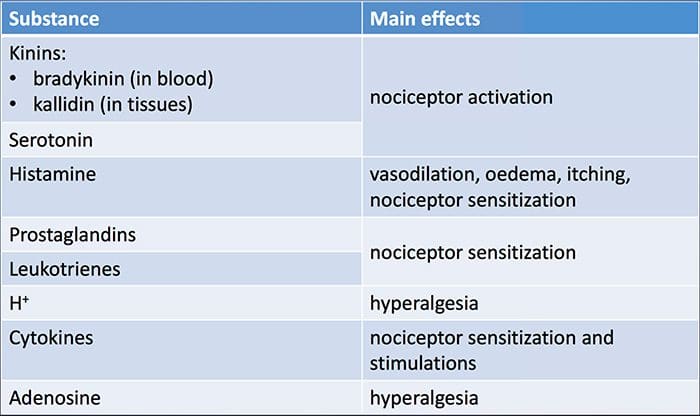
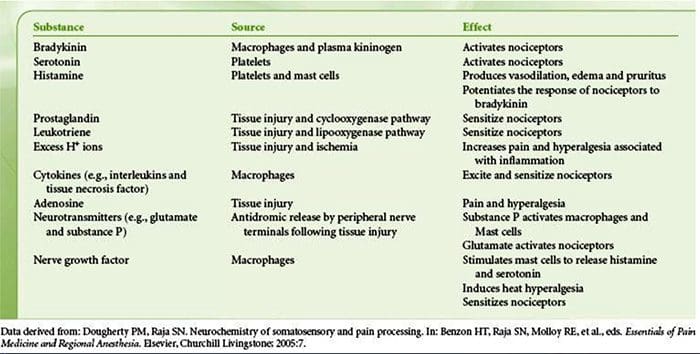
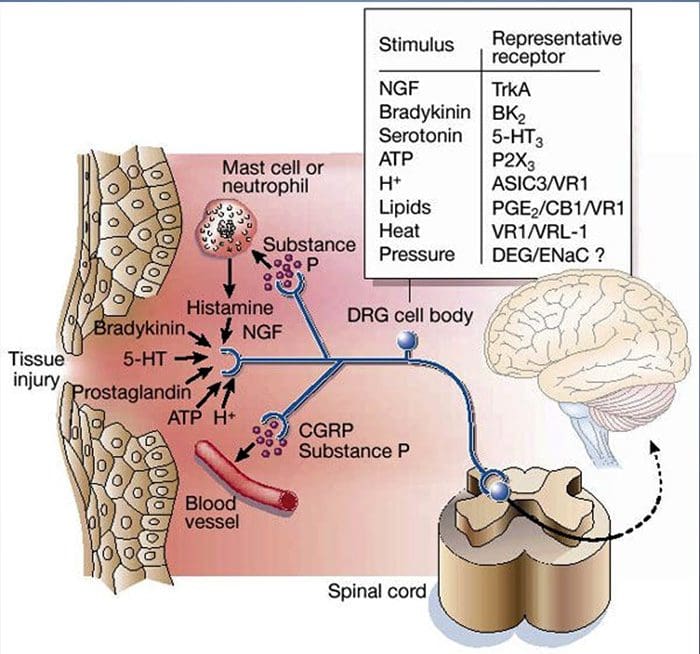
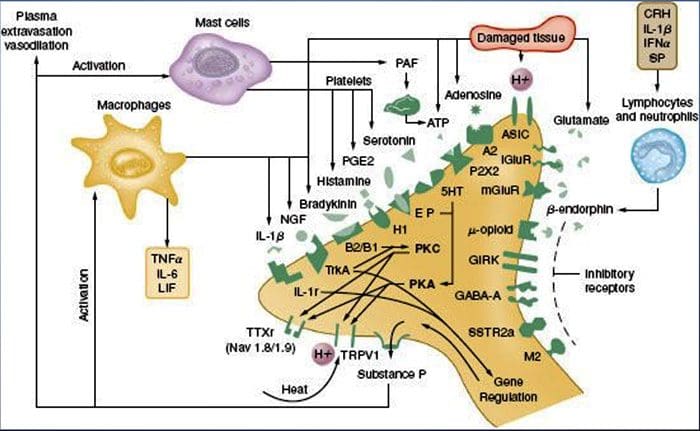 Perché mi fa male la spalla? Una rassegna delle basi neuroanatomiche e biochimiche del dolore alla spalla
Perché mi fa male la spalla? Una rassegna delle basi neuroanatomiche e biochimiche del dolore alla spalla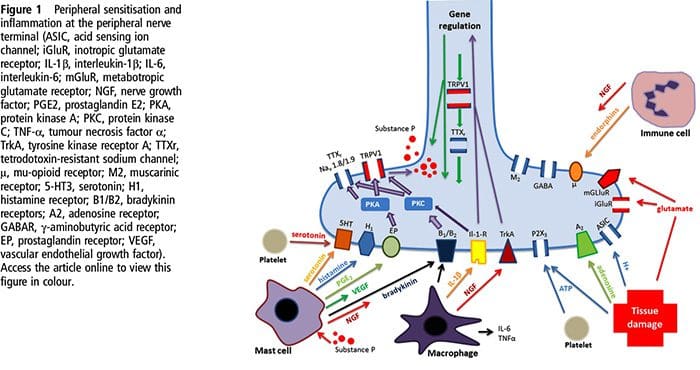 NGF e il recettore transitorio del potenziale recettore del canale cationico sottofamiglia V membro 1 (TRPV1) hanno una relazione simbiotica quando si tratta di infiammazione e sensibilizzazione dei nocicettori. Le citochine prodotte nel tessuto infiammato determinano un aumento della produzione di NGF.19 L'NGF stimola il rilascio di istamina e serotonina (5-HT3) da parte dei mastociti e sensibilizza anche i nocicettori, alterando eventualmente le proprietà di A? fibre tali che una percentuale maggiore diventa nocicettiva. Il recettore TRPV1 è presente in una sottopopolazione di fibre afferenti primarie ed è attivato da capsaicina, calore e protoni. Il recettore TRPV1 è sintetizzato nel corpo cellulare della fibra afferente e viene trasportato sia al terminale periferico che a quello centrale, dove contribuisce alla sensibilità delle afferenze nocicettive. L'infiammazione provoca la produzione periferica di NGF che si lega quindi al recettore della tirosin chinasi di tipo 1 sui terminali dei nocicettori, l'NGF viene quindi trasportato al corpo cellulare dove porta a una regolazione della trascrizione di TRPV1 e di conseguenza aumenta la sensibilità dei nocicettori.19 20 NGF e altri mediatori infiammatori sensibilizzano anche TRPV1 attraverso una vasta gamma di vie di messaggistica secondarie. Si ritiene che molti altri recettori, inclusi i recettori colinergici, i recettori dell'acido α-aminobutirrico (GABA) e i recettori della somatostatina, siano coinvolti nella sensibilità dei nocicettori periferici.
NGF e il recettore transitorio del potenziale recettore del canale cationico sottofamiglia V membro 1 (TRPV1) hanno una relazione simbiotica quando si tratta di infiammazione e sensibilizzazione dei nocicettori. Le citochine prodotte nel tessuto infiammato determinano un aumento della produzione di NGF.19 L'NGF stimola il rilascio di istamina e serotonina (5-HT3) da parte dei mastociti e sensibilizza anche i nocicettori, alterando eventualmente le proprietà di A? fibre tali che una percentuale maggiore diventa nocicettiva. Il recettore TRPV1 è presente in una sottopopolazione di fibre afferenti primarie ed è attivato da capsaicina, calore e protoni. Il recettore TRPV1 è sintetizzato nel corpo cellulare della fibra afferente e viene trasportato sia al terminale periferico che a quello centrale, dove contribuisce alla sensibilità delle afferenze nocicettive. L'infiammazione provoca la produzione periferica di NGF che si lega quindi al recettore della tirosin chinasi di tipo 1 sui terminali dei nocicettori, l'NGF viene quindi trasportato al corpo cellulare dove porta a una regolazione della trascrizione di TRPV1 e di conseguenza aumenta la sensibilità dei nocicettori.19 20 NGF e altri mediatori infiammatori sensibilizzano anche TRPV1 attraverso una vasta gamma di vie di messaggistica secondarie. Si ritiene che molti altri recettori, inclusi i recettori colinergici, i recettori dell'acido α-aminobutirrico (GABA) e i recettori della somatostatina, siano coinvolti nella sensibilità dei nocicettori periferici.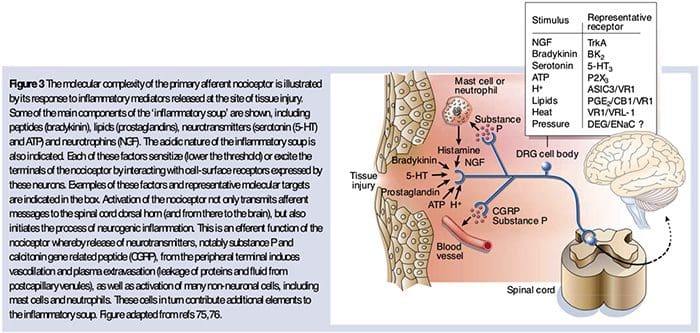 La neurochimica dei nocicettori
La neurochimica dei nocicettori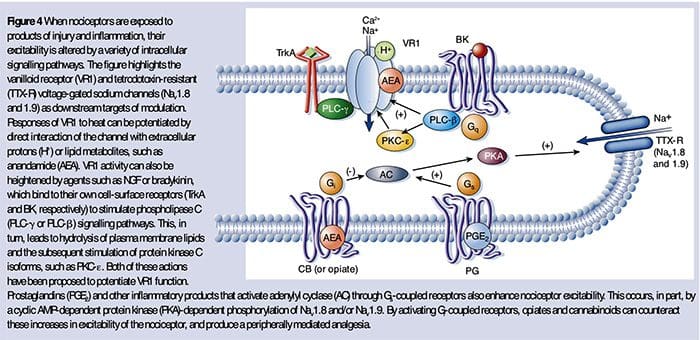 Meccanismi cellulari e molecolari del dolore
Meccanismi cellulari e molecolari del dolore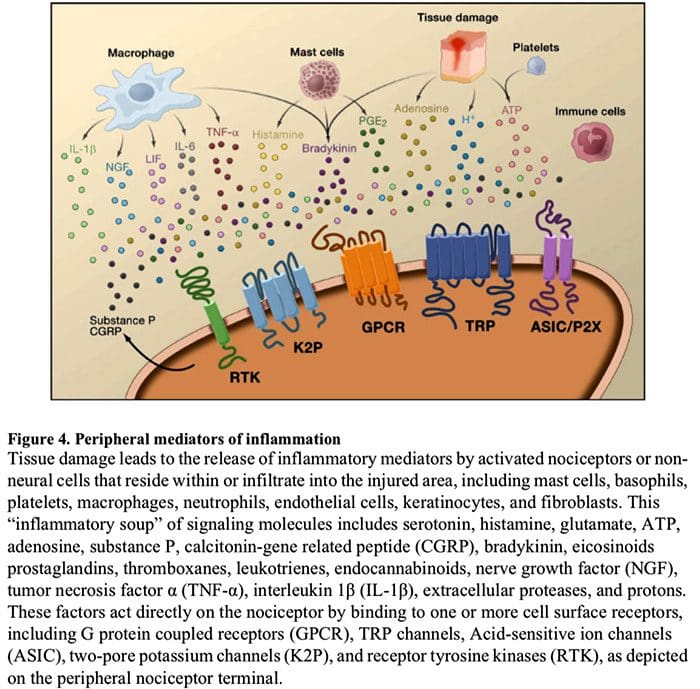
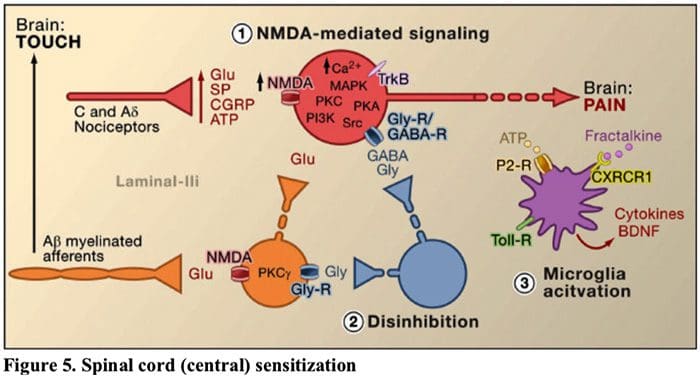 Figura 5. Sensibilizzazione al midollo spinale (centrale)
Figura 5. Sensibilizzazione al midollo spinale (centrale)