Supporto Clinico Neurofisiologico Back Clinic. El Paso, TX. Chiropratico, il Dott. Alexander Jimenez discute neurofisiologia clinica. Il dott. Jimenez esplorerà il significato clinico e le attività funzionali delle fibre nervose periferiche, del midollo spinale, del tronco cerebrale e del cervello nel contesto dei disturbi viscerali e muscolo-scheletrici. I pazienti acquisiranno una conoscenza avanzata dell'anatomia, della genetica, della biochimica e della fisiologia del dolore in relazione a varie sindromi cliniche. Verranno incorporati biochimica nutrizionale correlata alla nocicezione e al dolore. E l'applicazione di queste informazioni nei programmi di terapia sarà enfatizzata.
Il nostro team è molto orgoglioso di offrire alle nostre famiglie e ai pazienti feriti solo protocolli di trattamento comprovati. Insegnando il benessere olistico completo come stile di vita, cambiamo anche non solo la vita dei nostri pazienti, ma anche le loro famiglie. Lo facciamo in modo da poter raggiungere tutti gli El Pasoani che hanno bisogno di noi, indipendentemente dai problemi di accessibilità. Per le risposte a qualsiasi domanda tu possa avere, chiama il Dr. Jimenez al 915-850-0900.
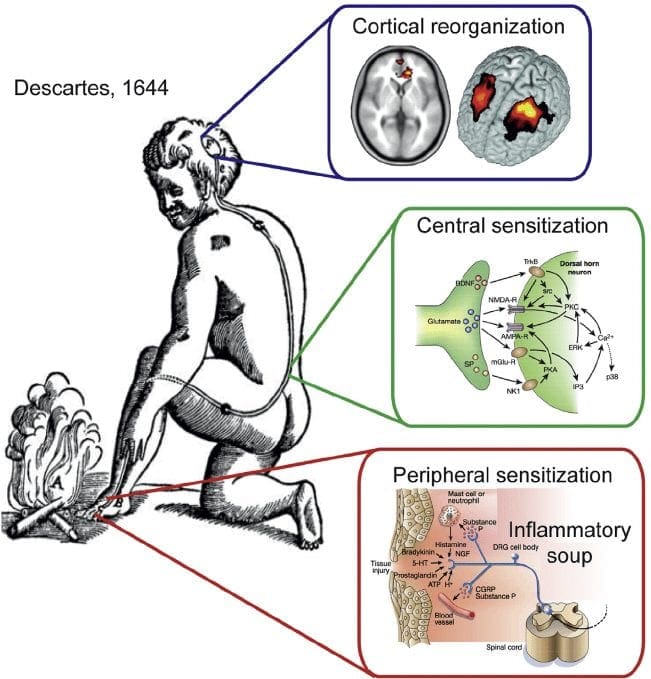
Il dolore neuropatico è una condizione complessa e cronica di dolore che è generalmente accompagnata da lesioni dei tessuti molli. Il dolore neuropatico è comune nella pratica clinica e rappresenta anche una sfida per pazienti e medici. Con il dolore neuropatico, le fibre nervose stesse possono essere danneggiate, disfunzionali o ferite. Il dolore neuropatico è il risultato di un danno da trauma o malattia al sistema nervoso periferico o centrale, in cui la lesione può verificarsi in qualsiasi sito. Di conseguenza, queste fibre nervose danneggiate possono inviare segnali errati ad altri centri di dolore. L'effetto di una lesione della fibra nervosa consiste in un cambiamento nella funzione neurale, sia nella regione della ferita che attorno alla lesione. I segni clinici del dolore neuropatico comprendono normalmente i fenomeni sensoriali, come il dolore spontaneo, le parestesie e l'iperalgesia.
Il dolore neuropatico, come definito dall'International Association of Study of Pain o dallo IASP, è il dolore iniziato o causato da una lesione primaria o disfunzione del sistema nervoso. Potrebbe derivare da danni ovunque lungo il neurotrasmettitore: sistema nervoso periferico, sistema nervoso spinale o sovraspinale. I tratti che distinguono il dolore neuropatico da altri tipi di dolore includono il dolore e i segni sensoriali che durano oltre il periodo di recupero. È caratterizzato negli esseri umani dal dolore spontaneo, dall'allodinia o dall'esperienza di stimolazione non nociva come dolorosa e causalgia o persistente dolore bruciante. Il dolore spontaneo include sensazioni di "spilli e aghi", bruciore, tiro, pugnalata e dolore parossistico, o shock elettrico come il dolore, spesso associato a disestesia e parestesie. Queste sensazioni non solo alterano l'apparato sensoriale del paziente, ma anche il benessere, l'umore, l'attenzione e il pensiero del paziente. Il dolore neuropatico è composto sia da sintomi "negativi", come perdita sensoriale e sensazioni di formicolio, sia da sintomi "positivi", come parestesie, dolore spontaneo e maggiore sensazione di dolore.
Le condizioni frequentemente correlate al dolore neuropatico possono essere classificate in due gruppi principali: dolore dovuto al danno nel sistema nervoso centrale e dolore a causa del danno al sistema nervoso periferico. Colpi corticali e sub-corticali, lesioni traumatiche del midollo spinale, siringo-myelia e siringobulbia, nevralgie del trigemino e glossofaringeo, neoplasie e altre lesioni che occupano spazio sono condizioni cliniche che appartengono al primo gruppo. Compressione nervosa o neuropatie intrappolate, neuropatia ischemica, polineuropatie periferiche, plexopatie, compressione delle radici nervose, moncone post-amputazione e dolore agli arti fantasma, nevralgia post-erpetica e neuropatie correlate al cancro sono condizioni cliniche che appartengono a quest'ultimo gruppo.
Fisiopatologia del dolore neuropatico
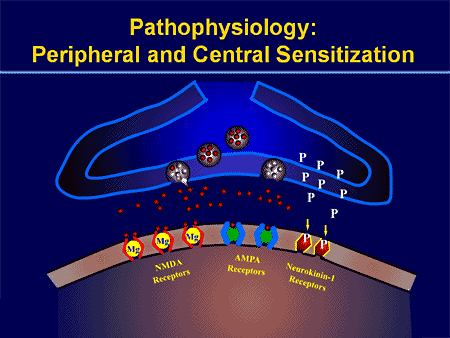
I processi e i concetti patofisiologici alla base del dolore neuropatico sono molteplici. Prima di coprire questi processi, una revisione dei circuiti del dolore ordinario è fondamentale. Regolari circuiti del dolore comportano l'attivazione di un nocicettore, noto anche come recettore del dolore, in risposta a una stimolazione dolorosa. Un'ondata di depolarizzazione viene trasmessa ai neuroni del primo ordine, insieme al sodio che scorre veloce attraverso i canali del sodio e il potassio che si sta esaurendo. I neuroni terminano nel tronco cerebrale nel nucleo del trigemino o nel corno dorsale del midollo spinale. È qui che il segno apre canali di calcio voltaggio-dipendenti nel terminale pre-sinaptico, consentendo l'ingresso di calcio. Il calcio consente al glutammato, un neurotrasmettitore eccitatorio, di essere rilasciato nell'area sinaptica. Il glutammato si lega ai recettori NMDA sui neuroni di secondo ordine, causando la depolarizzazione.
Questi neuroni attraversano il midollo spinale e viaggiano fino al talamo, dove si sintonizzano con i neuroni di terzo ordine. Questi si collegano quindi al sistema limbico e alla corteccia cerebrale. Esiste anche un percorso inibitorio che impedisce la trasmissione del segnale del dolore dal corno dorsale. I neuroni anti-nocicettivi hanno origine nel tronco cerebrale e viaggiano lungo il midollo spinale dove si sintetizzano con brevi interneuroni nel corno dorsale rilasciando dopamina e norepinefrina. Gli interneuroni modulano la sinapsi tra il neurone del primo ordine e il neurone del secondo ordine rilasciando acido gamma amino butirrico, o GABA, un neurotrasmettitore inibitorio. Di conseguenza, la cessazione del dolore è il risultato dell'inibizione delle sinapsi tra i neuroni del primo e del secondo ordine, mentre l'aumento del dolore potrebbe essere il risultato della soppressione delle connessioni sinaptiche inibitorie.
Il meccanismo alla base del dolore neuropatico, tuttavia, non è chiaro. Diversi studi su animali hanno rivelato che molti meccanismi possono essere coinvolti. Tuttavia, bisogna ricordare che ciò che si applica alle creature potrebbe non essere sempre applicabile alle persone. I neuroni del primo ordine possono aumentare il loro fuoco se sono parzialmente danneggiati e aumentare la quantità di canali del sodio. Gli scarichi ectopici sono una conseguenza della depolarizzazione potenziata in alcuni siti della fibra, con conseguente dolore spontaneo e dolore correlato al movimento. I circuiti inibitori potrebbero essere diminuiti nel livello del corno dorsale o delle cellule staminali del cervello, così come entrambi, permettendo agli impulsi del dolore di viaggiare senza opposizione.
Inoltre, potrebbero esserci delle alterazioni nell'elaborazione centrale del dolore quando, a causa del dolore cronico e dell'uso di alcuni farmaci e / o farmaci, i neuroni del secondo e terzo ordine possono creare una "memoria" del dolore e diventare sensibilizzati. C'è poi una maggiore sensibilità dei neuroni spinali e soglie di attivazione ridotte. Un'altra teoria dimostra il concetto di dolore neuropatico mantenuto in modo comprensivo. Questa nozione è stata dimostrata dall'analgesia in seguito a simpatectomia da parte di animali e persone. Tuttavia, un mix di meccanica può essere coinvolto in molte condizioni croniche neuropatiche o miste di dolore somatico e neuropatico. Tra quelle sfide nel campo del dolore, e molto più che riguarda il dolore neuropatico, c'è la capacità di controllarlo. C'è una duplice componente in questo: in primo luogo, valutare qualità, intensità e avanzamento; e in secondo luogo, diagnosticare correttamente il dolore neuropatico.
Esistono, tuttavia, alcuni strumenti diagnostici che possono aiutare i medici nella valutazione del dolore neuropatico. Per cominciare, studi sulla conduzione nervosa e potenziali evocati sensoriali possono identificare e quantificare l'entità del danno ai percorsi sensoriali, ma non nocicettivi, monitorando le risposte neurofisiologiche agli stimoli elettrici. Inoltre, il test sensoriale quantitativo misura la percezione in reazione a stimoli esterni di intensità variabile applicando la stimolazione alla pelle. La sensibilità meccanica agli stimoli tattili viene misurata con strumenti specializzati, come i peli di von Frey, la puntura a spillo con aghi a incastro, nonché la sensibilità alle vibrazioni insieme ai vibrametri e al dolore termico con i termodi.
È anche estremamente importante eseguire una valutazione neurologica completa per identificare le disfunzioni motorie, sensoriali e autonome. In definitiva, ci sono numerosi questionari usati per distinguere il dolore neuropatico nel dolore nocicettivo. Alcuni di essi includono solo domande di interviste (ad esempio, il Neuropathic Questionnaire e ID Pain), mentre altri contengono sia interviste che test fisici (ad esempio, la valutazione Leeds dei sintomi neuropatici e scala dei segni) e lo strumento romanzo esatto, la valutazione standardizzata di Dolore, che combina sei domande di intervista e dieci valutazioni fisiologiche.
Modalità di trattamento per il dolore neuropatico
I regimi farmacologici mirano ai meccanismi del dolore neuropatico. Tuttavia, entrambi i trattamenti farmacologici e non farmacologici forniscono sollievo completo o parziale in circa la metà dei pazienti. Molte testimonianze basate sull'evidenza suggeriscono l'uso di miscele di farmaci e / o farmaci per funzionare con il maggior numero possibile di meccanismi. La maggior parte degli studi ha studiato principalmente nevralgia post-erpetica e neuropatie diabetiche dolorose, ma i risultati potrebbero non essere applicabili a tutte le condizioni di dolore neuropatico.
antidepressivi
Gli antidepressivi aumentano i livelli di serotonina e norepinefrina sinaptica, migliorando così l'effetto del sistema analgesico discendente associato al dolore neuropatico. Sono stati il cardine della terapia del dolore neuropatico. Le azioni analgesiche potrebbero essere attribuibili al blocco del reuptake della nor-adrenalina e della dopamina, che presumibilmente aumentano l'inibizione discendente, l'antagonismo del recettore NMDA e il blocco del canale del sodio. Antidepressivi triciclici, come i TCA; ad esempio, amitriptilina, imipramina, nortriptilina e doxepina, sono potenti contro il dolore continuo e dolorante e il dolore spontaneo.
Gli antidepressivi triciclici si sono dimostrati significativamente più efficaci per il dolore neuropatico rispetto agli inibitori della ricaptazione della serotonina, o SSRI, come fluoxetina, paroxetina, sertralina e citalopram. La ragione potrebbe essere che inibiscono la ricaptazione di serotonina e nor-adrenalina, mentre gli SSRI inibiscono solo la ricaptazione della serotonina. Gli antidepressivi triciclici possono avere effetti collaterali spiacevoli, tra cui nausea, confusione, blocchi di conduzione cardiaca, tachicardia e aritmie ventricolari. Possono anche causare aumento di peso, una ridotta soglia convulsiva e ipotensione ortostatica. I triciclici devono essere usati con cautela negli anziani, che sono particolarmente vulnerabili ai loro effetti collaterali acuti. La concentrazione del farmaco nel sangue deve essere monitorata per evitare tossicità nei pazienti che sono metabolizzatori lenti del farmaco.
Gli inibitori del reuptake della serotonina e norepinefrina, o SNRI, sono una nuova classe di antidepressivi. Come i TCA, sembrano essere più efficaci degli SSRI per il trattamento del dolore neuropatico perché inibiscono anche la ricaptazione sia della nor-adrenalina che della dopamina. La venlafaxina è efficace contro la polineuropatia debilitante, come la neuropatia diabetica dolorosa, come l'imipramina, nella menzione del TCA, e i due sono significativamente maggiori del placebo. Come i TCA, gli SNRI sembrano conferire benefici indipendentemente dai loro effetti antidepressivi. Gli effetti collaterali includono sedazione, confusione, ipertensione e sindrome da astinenza.
Farmaci antiepilettici
I farmaci antiepilettici possono essere utilizzati come trattamento di prima linea soprattutto per alcuni tipi di dolore neuropatico. Agiscono modulando i canali voltaggio-dipendenti del calcio e del sodio, migliorando gli effetti inibitori del GABA e inibendo la trasmissione glutaminergica eccitatoria. Non è stato dimostrato che i farmaci antiepilettici siano efficaci per il dolore acuto. Nei casi di dolore cronico, i farmaci antiepilettici sembrano essere efficaci solo nella nevralgia del trigemino. La carbamazepina viene abitualmente utilizzata per questa condizione. Il gabapentin, che funziona inibendo la funzione del canale del calcio attraverso le azioni di agonista nella subunità delta alfa-2 del canale del calcio, è anche noto per essere efficace per il dolore neuropatico. Tuttavia, il gabapentin agisce a livello centrale e potrebbe causare affaticamento, confusione e sonnolenza.
Analgesici non oppioidi
Vi è una mancanza di dati forti a sostegno dell'uso di farmaci anti-infiammatori non steroidei o FANS, nel sollievo dal dolore neuropatico. Ciò può essere dovuto alla mancanza di un componente infiammatorio nell'alleviare il dolore. Ma sono stati utilizzati in modo intercambiabile con oppiacei come adiuvanti nel trattamento del dolore oncologico. Sono state segnalate complicazioni, tuttavia, specialmente in pazienti gravemente debilitati.
Analgesici oppioidi
Gli analgesici oppioidi sono oggetto di molti dibattiti nell'alleviare il dolore neuropatico. Agiscono inibendo gli impulsi del dolore ascendente centrale. Tradizionalmente, il dolore neuropatico è stato precedentemente osservato come resistente agli oppioidi, in cui gli oppioidi sono metodi più adatti per i tipi di dolore nocicettivo coronarico e somatico. Molti medici impediscono l'uso di oppiacei per il trattamento del dolore neuropatico, in gran parte a causa di preoccupazioni circa l'abuso di droghe, la dipendenza e le questioni normative. Ma ci sono molte prove che hanno trovato successo negli analgesici oppioidi. L'ossicodone era superiore al placebo per alleviare il dolore, l'allodinia, migliorare il sonno e l'handicap. Gli oppioidi a rilascio controllato, secondo una base programmata, sono raccomandati per i pazienti con dolore costante per incoraggiare livelli costanti di analgesia, prevenire fluttuazioni della glicemia e prevenire eventi avversi associati a dosaggi più elevati. Più comunemente, i preparati orali vengono utilizzati a causa della loro maggiore facilità d'uso ed economicità. I preparati trans-dermici, parenterali e rettali sono generalmente utilizzati in pazienti che non tollerano i farmaci per via orale.
Anestetici locali
Gli anestetici ad azione vicini sono allettanti perché, grazie alla loro azione regionale, hanno effetti collaterali minimi. Agiscono stabilizzando i canali del sodio agli assoni dei neuroni periferici di primo ordine. Funzionano meglio se sono presenti solo lesioni nervose parziali e sono stati rilevati canali di sodio in eccesso. La lidocaina topica è il rappresentante più studiato del corso per il dolore neuropatico. In particolare, l'uso di questo cerotto alla lidocaina 5 per la nevralgia post-erpetica ha causato la sua approvazione da parte della FDA. La patch sembra funzionare meglio quando è danneggiata, ma mantenuta, la funzione nocicettiva del sistema nervoso periferico dal dermatoma coinvolto che dimostra come allodinia. Deve essere impostato direttamente sull'area sintomatica per 12 ore ed eliminato per altre ore 12 e può essere utilizzato per anni in questo modo. Oltre alle reazioni cutanee locali, è spesso ben tollerato da molti pazienti con dolore neuropatico.
Droghe varie
La clonidina, un alfa-2-agonista, si è dimostrata efficace in un sottogruppo di pazienti con neuropatia periferica diabetica. È stato scoperto che i cannabinoidi svolgono un ruolo nella modulazione sperimentale del dolore in modelli animali e la prova dell'efficacia si sta accumulando. Gli agonisti selettivi di CB2 sopprimono iperalgesia e allodinia e normalizzano le soglie nocicettive senza indurre l'analgesia.
Gestione del dolore interventista
Trattamenti invasivi potrebbero essere presi in considerazione per i pazienti che hanno dolore neuropatico intrattabile. Questi trattamenti includono iniezioni epidurali o perineuriche di anestetici locali o corticosteroidi, l'impianto di metodi di somministrazione di farmaci epidurali e intratecali e l'inserimento di stimolatori del midollo spinale. Questi approcci sono riservati ai pazienti con dolore neuropatico cronico intrattabile che hanno fallito nella gestione medica conservativa e hanno anche sperimentato un'attenta valutazione psicologica. In uno studio di Kim et al, è stato dimostrato che uno stimolatore del midollo spinale era efficace nel trattamento del dolore neuropatico dell'origine delle radici nervose.
Insight di Dr. Alex Jimenez
Con il dolore neuropatico, i sintomi di dolore cronico si verificano a causa delle fibre nervose stesse danneggiate, disfunzionali o ferite, generalmente accompagnate da danni ai tessuti o lesioni. Di conseguenza, queste fibre nervose possono iniziare a inviare segnali dolorosi errati ad altre aree del corpo. Gli effetti del dolore neuropatico causato dalle lesioni delle fibre nervose comprendono modifiche nella funzione dei nervi sia nel sito di lesione che nelle aree attorno alla lesione. Comprendere la fisiopatologia del dolore neuropatico è stato un obiettivo per molti professionisti della salute, al fine di determinare in modo efficace l'approccio terapeutico migliore per aiutare a gestire e migliorare i sintomi. Dall'uso di droghe e / o farmaci, all'assistenza chiropratica, all'esercizio fisico, all'attività fisica e alla nutrizione, possono essere utilizzati una varietà di approcci terapeutici per alleviare il dolore neuropatico per i bisogni di ogni individuo.
Ulteriori interventi per il dolore neuropatico
Molti pazienti con dolore neuropatico perseguono opzioni di trattamento complementari e alternative per il trattamento del dolore neuropatico. Altri regimi ben noti usati per trattare il dolore neuropatico comprendono l'agopuntura, la stimolazione elettrica del nervo percutaneo, la stimolazione elettrica transcutanea del nervo, il trattamento cognitivo comportamentale, l'immaginazione motorio graduata e il trattamento di supporto e l'esercizio. Tra questi, tuttavia, la cura chiropratica è un noto approccio di trattamento alternativo comunemente usato per trattare il dolore neuropatico. La cura chiropratica, insieme alla fisioterapia, all'esercizio fisico, alla nutrizione e alle modifiche dello stile di vita possono alla fine offrire sollievo per i sintomi del dolore neuropatico.
Cura chiropratica
Ciò che è noto è che un'applicazione di gestione completa è fondamentale per combattere gli effetti del dolore neuropatico. In questo modo, la cura chiropratica è un programma di trattamento olistico che potrebbe essere efficace nel prevenire problemi di salute associati al danno ai nervi. La cura chiropratica fornisce assistenza a pazienti con molte condizioni diverse, comprese quelle con dolore neuropatico. Chi soffre di dolore neuropatico usa spesso farmaci anti-infiammatori non steroidei, o FANS, come l'ibuprofene, o antidolorifici pesanti per aiutare ad alleviare il dolore neuropatico. Questi possono fornire una soluzione temporanea, ma hanno bisogno di un uso costante per gestire il dolore. Ciò contribuisce invariabilmente a effetti collaterali dannosi e in situazioni estreme, dipendenza da farmaci da prescrizione.
La cura chiropratica può aiutare a migliorare i sintomi del dolore neuropatico e migliorare la stabilità senza questi aspetti negativi. Un approccio come la cura chiropratica offre un programma individualizzato progettato per individuare la causa principale del problema. Attraverso l'uso di regolazioni spinali e manipolazioni manuali, un chiropratico può correggere con cura qualsiasi disallineamento spinale, o sublussazioni, trovato lungo la lunghezza della colonna vertebrale, che potrebbe ridurre le conseguenze del nervo wracking attraverso il riallineamento della spina dorsale. Ripristinare l'integrità della colonna vertebrale è essenziale per mantenere un sistema nervoso centrale ad alto funzionamento.
Un chiropratico può anche essere un trattamento a lungo termine per migliorare il tuo benessere generale. Oltre agli aggiustamenti spinali e alle manipolazioni manuali, un chiropratico può offrire consigli nutrizionali, come prescrivere una dieta ricca di antiossidanti, oppure può progettare una terapia fisica o un programma di esercizi per combattere gli stimoli del dolore nervoso. Una condizione a lungo termine richiede un rimedio a lungo termine e, in questa veste, un operatore sanitario specializzato in lesioni e / o condizioni che colpiscono il sistema muscolo-scheletrico e nervoso, come un chiropratico o chiropratico, può essere prezioso durante il lavoro per valutare il cambiamento favorevole nel tempo.
È stato dimostrato che la terapia fisica, l'esercizio e le tecniche di rappresentazione del movimento sono benefiche per il trattamento del dolore neuropatico. La cura chiropratica offre anche altre modalità di trattamento che possono essere utili per la gestione o il miglioramento del dolore neuropatico. La terapia laser a basso livello, o LLLT, ad esempio, ha acquisito un'enorme importanza come trattamento per il dolore neuropatico. Secondo una serie di studi di ricerca, si è concluso che l'LLLT ha effetti positivi sul controllo dell'analgesia per il dolore neuropatico, tuttavia, sono necessari ulteriori studi per definire protocolli di trattamento che riassumano gli effetti della terapia laser a basso livello nei trattamenti neuropatici del dolore.
La cura chiropratica include anche consigli nutrizionali, che possono aiutare a controllare i sintomi associati alla neuropatia diabetica. Durante uno studio di ricerca è stata dimostrata una dieta a basso contenuto di grassi per migliorare il controllo glicemico nei pazienti con diabete tipo 2. Dopo circa 20 settimane dello studio pilota, le persone coinvolte riportarono cambiamenti nel loro peso corporeo e la conduttanza elettrochimica della pelle nel piede fu riportata come migliorata con l'intervento. Lo studio di ricerca ha suggerito un valore potenziale nell'intervento dietetico a base di grassi a base di piante per la neuropatia diabetica. Inoltre, studi clinici hanno scoperto che l'applicazione orale di L-treonato di magnesio è in grado di prevenire e ripristinare i deficit di memoria associati al dolore neuropatico.
La cura chiropratica può anche offrire strategie terapeutiche aggiuntive per promuovere la rigenerazione dei nervi. A titolo di esempio, è stato suggerito di migliorare la rigenerazione degli assoni per contribuire a migliorare il recupero funzionale dopo la lesione dei nervi periferici. Stimolazione elettrica, insieme con l'esercizio o attività fisica, è stata trovata per promuovere la rigenerazione dei nervi dopo la riparazione del nervo in ritardo negli esseri umani e nei ratti, secondo studi recenti. Alla fine, sia la stimolazione elettrica che l'esercizio fisico sono stati determinati come promettenti trattamenti sperimentali per la lesione dei nervi periferici che sembrano pronti per essere trasferiti all'uso clinico. Ulteriori studi di ricerca potrebbero essere necessari per determinare appieno gli effetti di questi nei pazienti con dolore neuropatico.
Conclusione
Il dolore neuropatico è un'entità multiforme senza particolari linee guida di cui prendersi cura. È meglio gestirlo utilizzando un approccio multidisciplinare. La gestione del dolore richiede una valutazione continua, l'educazione del paziente, la garanzia di follow-up e rassicurazione del paziente. Il dolore neuropatico è una condizione cronica che rende difficile scegliere il miglior trattamento. L'individualizzazione del trattamento implica la considerazione dell'impatto del dolore sul benessere, sulla depressione e sulle disabilità dell'individuo insieme alla formazione continua e alla valutazione. Gli studi sul dolore neuropatico, sia a livello molecolare che su modelli animali, sono relativamente nuovi ma molto promettenti. Sono previsti molti miglioramenti nei campi di base e clinici del dolore neuropatico, aprendo così le porte a modalità di trattamento migliori o nuove per questa condizione disabilitante. L'ambito delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
Depressione di ansia di doloreTutti hanno provato dolore, tuttavia, ci sono quelli con depressione, ansia o entrambi. Combina questo con il dolore e può diventare piuttosto intenso e difficile da trattare. Le persone che soffrono di depressione, ansia o entrambi tendono a provare dolore grave ea lungo termine più delle altre persone.
La via ansia, la depressione e il dolore si sovrappongono l'un l'altro è visto in sindromi croniche e in alcune sindromi dolorose invalidanti, cioè lombalgia, mal di testa, dolore nervoso e fibromialgia. I disturbi psichiatrici contribuiscono all'intensità del dolore e aumentano anche il rischio di disabilità.
Depressione:A (disturbo depressivo maggiore o depressione clinica) è un disturbo dell'umore comune ma grave. Provoca sintomi gravi che influenzano il modo in cui un individuo si sente, pensa e come gestisce le attività quotidiane, ad esempio dormire, mangiare e lavorare. Per essere diagnosticati con la depressione, i sintomi devono essere presenti per almeno due settimane.
Stato d'animo persistente, triste, ansioso o vuoto.
Sentimenti di disperazione, pessimisti.
Irritabilità.
Sentimenti di colpa, inutilità o impotenza.
Perdita di interesse o piacere nelle attività.
Diminuzione di energia o affaticamento.
Muoversi o parlare lentamente.
Sensazione di irrequietezza e difficoltà a stare seduti.
Difficoltà a concentrarsi, ricordare o prendere decisioni.
Difficoltà a dormire, risveglio mattutino e sonno eccessivo.
Appetito e cambiamenti di peso.
Pensieri di morte o suicidio e / o tentativi di suicidio.
Dolori, mal di testa, crampi o problemi digestivi senza una chiara causa fisica e / o che non si adattano al trattamento.
Non tutti coloro che sono depressi sperimentano ogni sintomo. Alcuni sperimentano solo pochi sintomi, mentre altri possono manifestarne diversi. Diversi sintomi persistenti oltre al cattivo umore sono necessarioper una diagnosi di depressione maggiore. La gravità e la frequenza dei sintomi insieme alla durata variano a seconda dell'individuo e della sua particolare malattia. I sintomi possono anche variare a seconda dello stadio della malattia.
DEPRESSIONE DI ANSIA DEL DOLORE
Obiettivi:
Qual è la relazione?
Cosa c'è dietro la neurofisiologia?
Quali sono le conseguenze principali?
Il cervello cambia nel dolore
Figura 1 Percorsi cerebrali, regioni e reti coinvolte nel dolore acuto e cronico
Davis, KD et al. (2017) Test di imaging cerebrale per il dolore cronico: problemi e raccomandazioni mediche, legali ed etiche Nat. Rev. Neurol. doi: 10.1038 / nrneurol.2017.122
DOLORE, ANSIA E DEPRESSIONE
Conclusione:
Il dolore, specialmente cronico, è associato a depressione e ansia
I meccanismi fisiologici che portano all'ansia e alla depressione possono essere di natura multifattoriale
Il dolore provoca cambiamenti in cervello struttura e funzione
Questo cambiamento nella struttura e nella funzione può alterare la capacità del cervello di modulare il dolore e controllare l'umore.
Quando il sistema sensoriale è colpito da lesioni o malattie, i nervi all'interno di quel sistema non possono funzionare correttamente per trasmettere sensazioni e sensazioni nel cervello. Questo spesso contribuisce ad una sensazione di intorpidimento o mancanza di sensibilità. Tuttavia, in alcuni casi, quando questo sistema è danneggiato, le persone possono provare dolore nell'area colpita.
Il dolore neuropatico non inizia bruscamente o si risolve rapidamente; è un il dolore cronico condizione che porta a sintomi dolorosi persistenti. Per la maggior parte delle persone, l'intensità dei loro sintomi può aumentare e diminuire durante il giorno. Anche se il dolore neuropatico dovrebbe essere correlato a problemi di salute dei nervi periferici, come la neuropatia causata da diabete o stenosi spinale, le lesioni al cervello o al midollo spinale possono anche portare a dolore neuropatico cronico. Il dolore neuropatico viene anche definito dolore nervoso.
Il dolore neuropatico può essere contrastato con il dolore nocicettivo. Il dolore neuropatico non si sviluppa in nessuna circostanza specifica o stimolo esterno, ma piuttosto i sintomi si verificano semplicemente perché il sistema nervoso potrebbe non funzionare di conseguenza. In effetti, gli individui possono anche provare dolore neuropatico anche quando la parte del corpo dolorante o ferita non è effettivamente lì. Questa condizione è chiamata dolore dell'arto fantasma, che può verificarsi nelle persone dopo che hanno avuto un'amputazione.
Il dolore nocicettivo è generalmente acuto e si sviluppa in risposta a una circostanza specifica, ad esempio quando qualcuno subisce un infortunio improvviso, come battere un dito con un martello o estirpare un dito quando si cammina a piedi nudi. Inoltre, il dolore nocicettivo tende ad andare via una volta guarito il sito interessato. Il corpo contiene cellule nervose specializzate, note come nocicettori, che rilevano stimoli nocivi che potrebbero danneggiare il corpo, come il caldo o il freddo estremo, la pressione, il pizzicamento e l'esposizione a sostanze chimiche. Questi segnali di avvertimento vengono quindi passati lungo il sistema nervoso al cervello, causando dolore nocicettivo.
Quali sono i fattori di rischio per il dolore neuropatico?
Qualsiasi cosa che contribuisca a una mancanza di funzione all'interno del sistema nervoso sensoriale può portare a dolore neuropatico. Pertanto, i problemi di salute dei nervi della sindrome del tunnel carpale o condizioni simili possono scatenare il dolore neuropatico. Il trauma, con conseguente lesione dei nervi, può portare a dolore neuropatico. Altre condizioni che potrebbero predisporre le persone a sviluppare il dolore neuropatico includono: diabete, carenze vitaminiche, cancro, HIV, ictus, sclerosi multipla, herpes zoster e persino alcuni trattamenti contro il cancro.
Quali sono le cause del dolore neuropatico?
Ci sono molte cause da cui gli individui possono sviluppare dolore neuropatico. Ma a livello cellulare, una spiegazione è un aumento del rilascio di alcuni recettori che indicano il dolore, insieme con una diminuita capacità dei nervi di modulare questi segnali, porta alla sensazione di dolore proveniente dalla regione colpita. Inoltre, nel midollo spinale, la regione che esercita segni dolorosi viene riorganizzata con corrispondenti cambiamenti negli ormoni e perdita di corpi mobili normalmente funzionanti. Quelle alterazioni provocano la percezione del dolore in assenza di stimolazione esterna. Nel cervello, la capacità di bloccare il dolore può essere influenzata da una lesione, come un ictus o un trauma. Col passare del tempo, si verifica un ulteriore danno cellulare e la sensazione di dolore continua. Il dolore neuropatico è anche correlato al diabete, all'assunzione cronica di alcol, a certi tumori, a carenza di vitamina B, a malattie, ad altre malattie correlate al sistema nervoso, a tossine e farmaci specifici.
Quali sono i sintomi del dolore neuropatico?
Contrariamente ad altre condizioni neurologiche, l'identificazione del dolore neuropatico può essere difficile. Tuttavia, possono essere presenti diversi segnali oggettivi. Gli operatori sanitari devono decifrare e tradurre un assortimento di parole che i pazienti usano per descrivere il loro dolore. I pazienti possono descrivere i loro sintomi come acuti, opachi, caldi, freddi, sensibili, pruriginosi, profondi, brucianti, tra una varietà di altri termini descrittivi. Inoltre, alcuni pazienti possono avvertire dolore attraverso il tocco leggero o la pressione.
Nello sforzo di aiutare a identificare la quantità di dolore che i pazienti potrebbero subire, vengono spesso utilizzate diverse scale. Ai pazienti viene chiesto di valutare il dolore secondo una scala visiva o un grafico numerico. Esistono molti esempi di scale del dolore, come quello illustrato di seguito. Spesso, le immagini di volti che raffigurano una varietà di livelli di dolore possono essere utili quando le persone hanno difficoltà a descrivere la quantità di dolore che stanno vivendo.
Dolore cronico e salute mentale
Per molti, l'impatto del dolore cronico non può essere limitato al dolore; può anche influenzare negativamente il loro stato mentale. Nuovi studi di ricerca condotti da scienziati della Northwestern University di Chicago possono spiegare perché le persone che soffrono di dolore cronico soffrono anche di problemi di salute apparentemente non correlati, come depressione, stress, mancanza di sonno e difficoltà di concentrazione.
La valutazione ha dimostrato che le persone con dolore cronico mostrano diverse regioni del cervello che sono sempre attive, in particolare l'area associata a umore e attenzione. Questa azione continua rimuove le connessioni nervose dal cervello e lascia chi soffre di dolore cronico a maggior rischio di problemi psicologici. I ricercatori hanno suggerito che ottenere costantemente segnali di dolore potrebbe comportare un ricablaggio mentale che influisce negativamente sulla mente. Il ricablaggio costringe il loro cervello a dedicare risorse mentali in modo diverso per affrontare le attività quotidiane, dalla matematica, al richiamo di una lista della spesa, al sentirsi felici.
La connessione dolore-cervello è stata ben registrata, almeno aneddoticamente, e molti operatori sanitari affermano di aver visto in prima persona il modo in cui lo stato mentale del paziente può andare in discesa quando sopportano il dolore cronico. Fraintendimenti circa la connessione dolore-cervello possono essere emersi dalla mancanza di prove che il dolore ha un'influenza misurabile e duratura sul cervello. I ricercatori si aspettano che, con ulteriori ricerche sui meccanismi di come il dolore cronico rende le persone più suscettibili ai disturbi dell'umore, le persone avranno la capacità di gestire meglio il loro benessere generale.
Cultura e dolore cronico
Molte cose contribuiscono al modo in cui sperimentiamo ed esprimiamo dolore, tuttavia, è stato anche recentemente suggerito dai ricercatori che la cultura si rapporta direttamente all'espressione del dolore. La nostra educazione e valori sociali influenzano il modo in cui esprimiamo il dolore e anche la sua natura, intensità e lunghezza. Tuttavia, queste variabili non sono così ovvie come i valori socio-psicologici, come l'età e il sesso.
La ricerca afferma che il dolore cronico è un processo multiforme e la concomitante interazione tra fattori fisiopatologici, cognitivi, affettivi, comportamentali e socioculturali riassume ciò che viene definito come l'esperienza del dolore cronico. È emerso che il dolore cronico è vissuto in modo diverso tra i pazienti di varie culture ed etnie.
Alcune culture incoraggiano l'espressione del dolore, in particolare nel Mediterraneo meridionale e in Medio Oriente. Altre persone lo sopprimono, come nelle molte lezioni ai nostri bambini sul comportarsi coraggiosamente e non piangere. Il dolore è riconosciuto come parte dell'esperienza umana. Siamo propensi ad assumere che la comunicazione sul dolore attraversi senza soluzione di continuità i confini culturali. Ma le persone che soffrono sono soggette ai modi in cui le loro civiltà li hanno addestrati a sperimentare ed esprimere dolore.
Entrambe le persone nel dolore e gli operatori sanitari hanno difficoltà a comunicare il dolore oltre i confini etnici. In una questione come il dolore, in cui una comunicazione efficace può avere implicazioni di vasta portata per l'assistenza medica, la qualità della vita e potenzialmente la sopravvivenza, il ruolo della cultura nella comunicazione del dolore rimane sottovalutato. Il dolore persistente è un incontro multidimensionale, composito formato da intrecci e co-influenzanti fattori biologici e psicosociali. Conoscere il culmine di questi fattori è fondamentale per comprendere le differenze della sua manifestazione e gestione.
Come viene diagnosticato il dolore neuropatico?
La diagnosi del dolore neuropatico si basa sulla valutazione aggiuntiva della storia di un individuo. Se si sospetta un danno nervoso sottostante, l'analisi dei nervi e il test possono essere giustificati. Il mezzo più comune per valutare se un nervo è ferito o meno sta usando la medicina elettrodiagnostica. Questa sottospecializzazione medica utilizza tecniche di studi di conduzione nervosa con elettromielografia (NCS / EMG). La valutazione clinica può mostrare evidenza di perdita di lavoro e può includere la valutazione del tocco leggero, la capacità di differenziare il dolore sordo e la capacità di discernere la temperatura, così come la valutazione della vibrazione.
Dopo aver completato un esame clinico approfondito, è possibile pianificare l'analisi elettrodiagnostica. Questi studi sono condotti da neurologi e fisiatri appositamente formati. Se si sospetta la neuropatia, una ricerca per cause reversibili dovrebbe essere compiuta. Questo può includere la funzione del sangue per carenze vitaminiche o problemi alla tiroide e studi di imaging per escludere una lesione strutturale che interessa il midollo spinale. A seconda dei risultati di questo test, potrebbe esserci un mezzo per diminuire l'intensità della neuropatia e possibilmente ridurre il dolore che un paziente sta vivendo.
Purtroppo, in molte condizioni, persino un buon controllo della causa sottostante della neuropatia non può invertire il dolore neuropatico. Questo è comunemente visto in pazienti con neuropatia diabetica. In rari casi, ci possono essere segni di cambiamenti nel modello di crescita della pelle e dei capelli in una regione colpita. Queste alterazioni possono essere associate a cambiamenti nel sudore. Se presenti, questi cambiamenti possono aiutare a identificare la probabile presenza di dolore neuropatico correlato a una condizione nota come sindrome da dolore regionale complesso.
Insight di Dr. Alex Jimenez
Il dolore neuropatico è una condizione di dolore cronico che è generalmente associata a danni diretti o lesioni al sistema nervoso o ai nervi. Questo tipo di dolore è diverso dal dolore nocicettivo o dalla tipica sensazione di dolore. Il dolore nocicettivo è una sensazione acuta o improvvisa di dolore che induce il sistema nervoso a inviare segnali di dolore subito dopo il trauma. Con il dolore neuropatico, tuttavia, i pazienti possono provare a sparare, bruciando il dolore senza alcun danno diretto o lesioni. Comprendere le possibili cause del dolore neuropatico del paziente rispetto a qualsiasi altro tipo di dolore, può aiutare gli operatori sanitari a trovare modi migliori per trattare le condizioni di dolore cronico.
Qual è il trattamento per il dolore neuropatico?
Vari farmaci sono usati nel tentativo di trattare il dolore neuropatico. La maggior parte di questi farmaci sono utilizzati off-label, il che significa che il farmaco è stato approvato dalla FDA per trattare diverse condizioni ed è stato quindi riconosciuto come vantaggioso per il trattamento del dolore neuropatico. Gli antidepressivi triciclici, come l'amitriptilina, la nortriptilina e la desipramina, sono stati prescritti per la gestione del dolore neuropatico per diversi anni.
Alcuni individui ritengono che questi possano essere molto efficaci nel dare loro sollievo. Altri tipi di antidepressivi hanno dimostrato di offrire sollievo. In alcuni pazienti sono stati utilizzati inibitori selettivi della ricaptazione della serotonina, o SSRI, come paroxetina e citalopram e altri antidepressivi, come la venlafaxina e il bupropione. Un altro trattamento frequente del dolore neuropatico comprende farmaci anti-aisenzici, tra cui carbamazepina, fenitoina, gabapentin, lamotrigina e altri.
Nei casi acuti di neuropatia dolorosa che non rispondono ai broker di prima linea, i farmaci tipicamente utilizzati per trattare le aritmie cardiache possono essere di qualche beneficio; tuttavia, questi possono portare a significativi effetti collaterali e spesso devono essere monitorati attentamente. I farmaci applicati direttamente sulla pelle possono offrire un beneficio modesto a quello percepibile per alcuni pazienti. Le forme comunemente usate includono la lidocaina (in patch o in gel) o la capsaicina.
Il trattamento del dolore neuropatico dipende dalla causa sottostante. Se la causa è reversibile, i nervi periferici possono rigenerarsi e il dolore diminuirà; tuttavia, questa riduzione del dolore può richiedere diversi mesi o anni. Molte altre opzioni di trattamento alternative, tra cui la cura chiropratica e la terapia fisica, possono anche essere utilizzate al fine di alleviare la tensione e lo stress lungo i nervi, contribuendo in definitiva a migliorare i sintomi dolorosi.
Qual è la prognosi per il dolore neuropatico?
Molte persone con dolore neuropatico sono in grado di ottenere qualche misura di aiuto, anche quando il loro dolore persiste. Sebbene il dolore neuropatico non sia pericoloso per un paziente, la presenza di dolore cronico può influire negativamente sulla qualità della vita. I pazienti con dolore cronico ai nervi potrebbero soffrire di privazione del sonno o disturbi dell'umore, inclusi depressione, ansia e stress, come precedentemente menzionato sopra. A causa dell'alopecia intrinseca e della mancanza di feedback sensoriale, i pazienti sono a rischio di sviluppare lesioni o infezioni o causare inconsapevolmente un'escalation di una lesione presente. Pertanto, è essenziale consultare immediatamente un medico e seguire le linee guida specifiche dirette da un operatore sanitario per sicurezza e prudenza.
Il dolore neuropatico può essere prevenuto?
Il modo migliore per prevenire il dolore neuropatico è evitare lo sviluppo o la progressione della neuropatia. Monitoraggio e modifica delle opzioni di stile di vita, inclusa la limitazione dell'uso di alcol e tabacco; mantenere un peso sano per ridurre la possibilità di diabete, malattie degenerative delle articolazioni o ictus; e avere una grande forma ergonomica al lavoro o quando si praticano hobby per ridurre il rischio di lesioni da stress ripetitivo sono strategie per diminuire la probabilità di sviluppare neuropatia e potenziale dolore neuropatico. Assicurati di consultare immediatamente un medico in caso di sintomi associati al dolore neuropatico al fine di procedere con l'approccio terapeutico più appropriato.L'ambito delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
Se il sistema sensoriale viene influenzato da lesioni o malattie, i nervi in quel sistema non possono funzionare nella trasmissione delle sensazioni al cervello. Questo può portare a una sensazione di intorpidimento o mancanza di sensibilità. In alcuni casi, quando il sistema sensoriale è danneggiato, le persone possono provare dolore nella regione interessata. Il dolore neuropatico non inizia rapidamente o termina rapidamente. È una condizione cronica che porta a sintomi di dolore persistente. Per molti, l'intensità dei sintomi può andare e venire per tutto il giorno. Si ritiene che il dolore neuropatico sia associato a problemi dei nervi periferici, cioè la neuropatia causata da diabete, stenosi spinale, lesioni al cervello o al midollo spinale possono anche portare a dolore neuropatico cronico.
DOLORE NEUROPATICO
Obiettivi:
Che cos'è?
Qual è la fisiopatologia che c'è dietro?
Quali sono le cause
Quali sono alcuni dei percorsi
Come possiamo aggiustarlo?
DOLORE NEUROPATICO
Dolore iniziato o causato da una lesione o disfunzione primaria nel sistema nervoso somatosensoriale.
Il dolore neuropatico è solitamente cronico, difficile da trattare e spesso resistente alla normale gestione analgesica.
PATOGENESI DEL DOLORE NEUROPATICO
MECCANISMI PERIFERICI
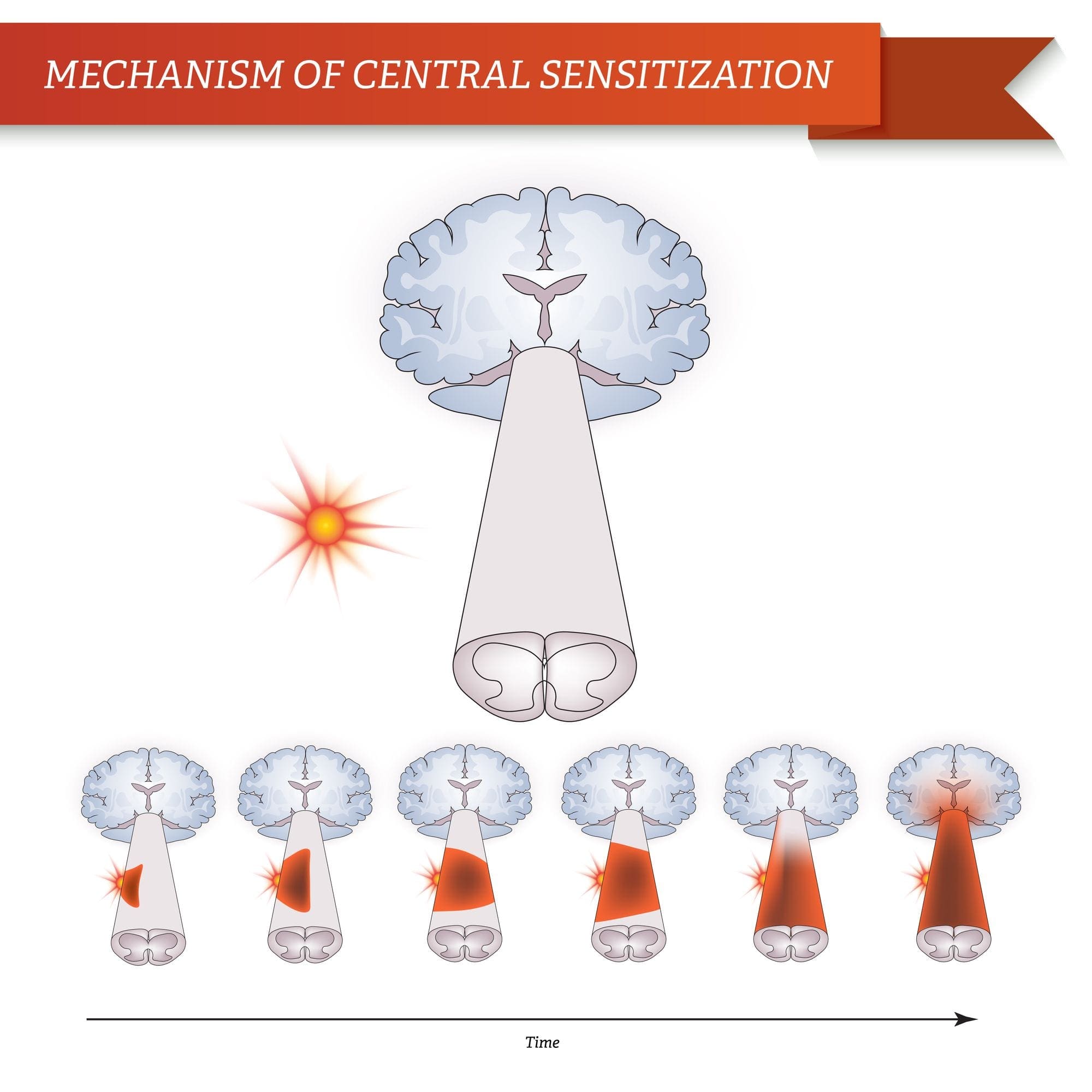
Dopo una lesione del nervo periferico, i neuroni diventano più sensibili e sviluppano un'eccitabilità anomala e un'elevata sensibilità alla stimolazione
Questo è noto come ... Sensibilizzazione periferica!
MECCANISMI CENTRALI
Come conseguenza della continua attività spontanea che si manifesta nella periferia, i neuroni sviluppano una maggiore attività di fondo, campi di recettività allargati e maggiori risposte agli impulsi afferenti, compresi i normali stimoli tattili
Questo è noto come ...Sensibilizzazione centrale!
CAUSE COMUNI
Lesioni o malattie del sistema nervoso somatosensoriale possono portare a una trasmissione alterata e disordinata di segnali sensoriali nel midollo spinale e nel cervello; le condizioni comuni associate al dolore neuropatico includono:
Nevralgia posterpetica
Nevralgia del trigemino
Radicolopatia dolorosa
Neuropatia diabetica
Infezione da HIV
Lebbra
Amputazione
Dolore alle lesioni nervose periferiche
Ictus (sotto forma di dolore post-ictus centrale)
DOLORE DEGLI ARTI FANTASMA E REALTÀ AUMENTATA
Phantom Limb Pain e AR
INFIAMMAZIONE NEUROGENICA
Obiettivi:
Che cos'è?
Qual è la fisiopatologia che c'è dietro?
Quali sono le cause
Come possiamo aggiustarlo?
INFIAMMAZIONE NEUROGENICA
Infiammazione neurogena è una risposta infiammatoria localizzata neurologicamente, caratterizzata da vasodilatazione, aumento della permeabilità vascolare, degranulazione dei mastociti e rilascio di neuropeptidi incluso il peptide correlato al gene della SP e della calcitonina (CGRP)
Sembra giocare un ruolo importante nella patogenesi di numerose malattie tra cui emicrania, psoriasi, asma, fibromialgia, eczema, rosacea, distonia e sensibilità chimica multipla
CAUSE COMUNI
Esistono molteplici vie attraverso le quali può essere avviata l'infiammazione neurogena. E 'ben documentato, utilizzando sia modelli animali che neuroni isolati in vitro, che la capsaicina, il calore, i protoni, la bradichinina e la triptasi sono regolatori a monte dell'influenza del calcio intracellulare, che provoca il rilascio di neuropeptidi infiammatori. Al contrario, si ritiene che le prostaglandine E2 e I2, citochine, interleuchina-1, interleuchina-6 e fattore di necrosi tumorale non causino il rilascio dei neurotrasmettitori stessi, ma piuttosto eccitino i neuroni sensoriali e quindi abbassino la soglia per sparare e causino il rilascio accresciuto di neuropeptidi.
Mentre l'infiammazione neurogena è stata ampiamente studiata e ben documentata nei tessuti periferici, fino a poco tempo fa il concetto di infiammazione neurogena all'interno del sistema nervoso centrale era rimasto in gran parte inesplorato. Data la capacità dell'infiammazione neurogena di influenzare la permeabilità vascolare e portare alla genesi dell'edema, è stato ora ampiamente studiato per il suo potenziale di influenzare la permeabilità del BBB e l'edema vasogenico nel cervello e nel midollo spinale in varie condizioni patologiche.
Siamo stati tutti educati da bambini che ci sono i sensi 5: vista, gusto, suono, odore e tatto. I quattro sensi iniziali utilizzano organi chiari e distinti, come gli occhi, le papille gustative, le orecchie e il naso, ma come fa il corpo a percepire il tocco esattamente? Il tatto è vissuto su tutto il corpo, sia all'interno che all'esterno. Non esiste un organo distinto responsabile del tocco sensibile. Piuttosto, ci sono piccoli recettori o terminazioni nervose attorno a tutto il corpo che percepiscono il contatto dove si verifica e invia segnali al cervello con informazioni sul tipo di tocco che si è verificato. Come un germoglio del gusto sulla lingua rileva il sapore, i meccanorecettori sono ghiandole all'interno della pelle e su altri organi che rilevano sensazioni tattili. Sono conosciuti come meccanorecettori perché sono progettati per rilevare sensazioni meccaniche o differenze di pressione.
Ruolo dei meccanocettori
Una persona comprende di aver provato una sensazione quando l'organo responsabile della scoperta di quel senso specifico invia un messaggio al cervello, che è l'organo principale che elabora e organizza tutte le informazioni. I messaggi vengono inviati da tutte le aree del corpo al cervello attraverso i fili indicati come neuroni. Ci sono migliaia di piccoli neuroni che si diramano verso tutte le aree del corpo umano, e sulle terminazioni di molti di questi neuroni sono i meccanorecettori. Per dimostrare cosa succede quando tocchi un oggetto, useremo un esempio.
Immagina una zanzara atterra sul tuo braccio. Il ceppo di questo insetto, così leggero, stimola i meccanocettori in quella particolare area del braccio. Quei meccanorecettori mandano un messaggio lungo il neurone a cui sono collegati. Il neurone si connette fino al cervello, che riceve il messaggio che qualcosa sta toccando il tuo corpo nella posizione esatta dello specifico meccanorecettore che ha inviato il messaggio. Il cervello agirà con questo consiglio. Forse dirà agli occhi di guardare la regione del braccio che ha rilevato la firma. E quando gli occhi dicono al cervello che c'è una zanzara sul braccio, il cervello potrebbe dire alla mano di spostarla rapidamente. Ecco come funzionano i meccanocettori. Lo scopo di questo articolo è dimostrare e discutere in dettaglio l'organizzazione funzionale e i determinanti molecolari dei meccanocettori.
Senso tattile: organizzazione funzionale e determinanti molecolari dei recettori meccanosensibili
Astratto
I meccanocettori cutanei sono localizzati nei vari strati della pelle dove rilevano un'ampia gamma di stimoli meccanici, tra cui pennello leggero, stiramento, vibrazione e pressione nociva. Questa varietà di stimoli è accompagnata da una vasta gamma di meccanorecettori specializzati che rispondono alla deformazione cutanea in un modo specifico e trasmettono questi stimoli a strutture cerebrali più elevate. Gli studi sui meccanocettori e sulle terminazioni nervose sensoriali geneticamente tracciabili stanno iniziando a scoprire i meccanismi della sensazione tattile. Il lavoro in questo campo ha fornito ai ricercatori una comprensione più approfondita dell'organizzazione del circuito alla base della percezione del tatto. Nuovi canali ionici sono emersi come candidati per le molecole di trasduzione e le proprietà delle correnti a controllo meccanico migliorano la nostra comprensione dei meccanismi di adattamento agli stimoli tattili. Questa recensione mette in evidenza i progressi compiuti nella caratterizzazione delle proprietà funzionali dei meccanocettori in cellule pelose e glabre e canali ionici che rilevano input meccanici e modellano l'adattamento meccanocettori.
parole chiave:meccanorecettore, canale meccanosensitivo, dolore, pelle, sistema somatosensoriale, tatto
Introduzione
Il tocco è la rilevazione di stimoli meccanici che colpiscono la pelle, inclusi stimoli meccanici innocui e nocivi. È un senso essenziale per la sopravvivenza e lo sviluppo di mammiferi e umani. Il contatto di oggetti solidi e fluidi con la pelle fornisce le informazioni necessarie al sistema nervoso centrale che consente l'esplorazione e il riconoscimento dell'ambiente e avvia la locomozione o il movimento pianificato della mano. Il tocco è anche molto importante per l'apprendistato, i contatti sociali e la sessualità. Il senso del tatto è il senso meno vulnerabile, sebbene possa essere distorto (iperestesia, ipoestesia) in molte condizioni patologiche.1-3
Le risposte al tocco implicano una codifica molto precisa delle informazioni meccaniche. I meccanocettori cutanei sono localizzati nei vari strati della pelle dove rilevano un'ampia gamma di stimoli meccanici, tra cui pennello leggero, allungamento, vibrazione, deflessione dei capelli e pressione nociva. Questa varietà di stimoli è abbinata a una vasta gamma di meccanocettori specializzati che rispondono alla deformazione cutanea in un modo specifico e trasmettono questi stimoli a strutture cerebrali superiori. I neuroni somatosensoriali della pelle si dividono in due gruppi: meccanocettori a bassa soglia (LTMR) che reagiscono alla pressione benigna e meccanocettori ad alta soglia (HTMR) che rispondono alla stimolazione meccanica dannosa. I corpi cellulari LTMR e HTMR risiedono all'interno dei gangli della radice dorsale (DRG) e dei gangli sensoriali cranici (gangli del trigemino). Le fibre nervose associate a LTMR e HTMR sono classificate come fibre A? -, A? - o fibre C in base alle loro velocità di conduzione del potenziale d'azione. Le fibre C sono non mielinizzate e hanno le velocità di conduzione più lente (~ 2 m / s), mentre A? e A? le fibre sono leggermente e pesantemente mielinizzate, mostrando rispettivamente velocità di conduzione intermedie (~ 12 m / s) e rapide (~ 20 m / s). Le LTMR sono anche classificate come risposte ad adattamento lento o rapido (SA- e RA-LTMR) in base alle loro velocità di adattamento allo stimolo meccanico sostenuto. Si distinguono inoltre per gli organi terminali cutanei che innervano e per i loro stimoli preferiti.
La capacità dei meccanocettori di rilevare i segnali meccanici si basa sulla presenza di canali ionici a trasduttore di velocità che trasformano rapidamente le forze meccaniche in segnali elettrici e depolarizzano il campo ricettivo. Questa depolarizzazione locale, chiamata potenziale recettore, può generare potenziali d'azione che si propagano verso il sistema nervoso centrale. Tuttavia, le proprietà delle molecole che mediano la meccanotrasduzione e l'adattamento alle forze meccaniche rimangono poco chiare.
In questa recensione, forniamo una panoramica delle proprietà dei meccanorecettori dei mammiferi nel tocco innocuo e nocivo nella pelle pelosa e glabra. Consideriamo anche le recenti conoscenze sulle proprietà delle correnti meccanicamente controllate nel tentativo di spiegare il meccanismo di adattamento dei meccanorecettori. Infine, esaminiamo i recenti progressi compiuti nell'identificazione dei canali ionici e delle proteine associate responsabili della generazione di correnti meccanizzate.
Tocco innocente
LTMR associati ai follicoli piliferi
I follicoli piliferi rappresentano i mini-organi che producono l'albero dei capelli che rilevano un tocco leggero. Le fibre associate ai follicoli piliferi rispondono al movimento dei capelli e alla loro direzione facendo scoppiare treni di potenziali d'azione all'inizio e alla rimozione dello stimolo. Stanno adattando rapidamente i recettori.
Gatto e coniglio Nel pelo di gatto e coniglio, i follicoli piliferi possono essere suddivisi in tre tipi di follicoli piliferi, i capelli in piuma, i capelli di guardia e i Tylotrichs. I peli Down (underhair, wool, vellus) 4 sono i peli più numerosi, i più corti e più fini del mantello. Sono ondulati, incolori e sono emersi in gruppi da due a quattro peli da un orifizio comune nella pelle. I peli della Guardia (monotrich, orecchio, tophair) 4 sono leggermente ricurvi, pigmentati o non pigmentati, ed emergono singolarmente dalla bocca dei loro follicoli. I tylotrich sono i peli meno numerosi, i più lunghi e più spessi.5,6 Sono pigmentati o non pigmentati, a volte entrambi ed emergono singolarmente da un follicolo circondato da un'ansa di vasi sanguigni capillari. Le fibre sensoriali fornite a un follicolo pilifero si trovano sotto la ghiandola sebacea e sono attribuite ad A? o fibre A? -LTMR 7
In stretta aderenza al fusto dei capelli, appena sotto il livello della ghiandola sebacea si trova l'anello delle terminazioni pilo-Ruffini lanceolate. Queste terminazioni nervose sensoriali sono posizionate in un percorso a spirale attorno al fusto del capello all'interno del tessuto connettivo che forma il follicolo pilifero. All'interno del follicolo pilifero ci sono anche terminazioni nervose libere, alcune delle quali formano meccanocettori. Frequentemente, i corpuscoli tattili (vedi pelle glabra) circondano la regione del collo del follicolo tylotrich.
Le proprietà delle terminazioni nervose mieliniche nella pelle pelosa di gatto e coniglio sono state esplorate intensamente nel periodo 1930-1970 (revisione in Hamann, 1995) .8 Notevolmente, Brown e Iggo, studiando 772 unità con fibre nervose afferenti mielinizzate nei nervi safeni di gatto e coniglio, hanno classificato le risposte in tre tipi di recettori corrispondenti ai movimenti dei peli di Down (recettori di tipo D), dei capelli di Guard (recettori di tipo G) e dei capelli di Tylotrich (recettori di tipo T) .9 Tutte le risposte delle fibre nervose afferenti sono state riunite nel recettore ad adattamento rapido di tipo I (RA I) per opposizione al recettore paciniano denominato RA II. I meccanocettori RA I rilevano la velocità dello stimolo meccanico e hanno un bordo netto. Non rilevano variazioni termiche. Burgess et al. ha anche descritto un recettore di campo ad adattamento rapido che risponde in modo ottimale allo sfregamento della pelle o al movimento di più peli, attribuito alla stimolazione delle terminazioni pilo-Ruffini. Nessuna risposta del follicolo pilifero è stata attribuita all'attività della fibra C. 10
Topi. Nella dorsale pelle pelosa di topi, sono stati descritti tre principali tipi di follicoli piliferi. Zigzag (circa 72%), punteruolo / auchene (circa 23%) e la guardia o tylotrich (circa 5%) 11-14 Zigzag e punteruolo / i follicoli piliferi auchenne producono i fusti dei capelli più sottili e più corti e sono associati a una ghiandola sebacea. I peli di guardia o tylotrich sono i più lunghi tra i tipi di follicoli piliferi. Sono caratterizzati da un bulbo pilifero di grandi dimensioni associato a due ghiandole sebacee. I peli di guardia e pelo / auchene sono disposti in uno schema iterativo, regolarmente distanziato, mentre i peli a zigzag popolano densamente le aree della pelle che circondano i due tipi di follicolo pilifero più grandi [Fig. 1 (A1, A2 e A3)].
Immagine 1. Organizzazione e proiezioni di meccanocettori cutanei. Nella pelle pelosa, il pennello leggero e il tatto sono rilevati principalmente dall'innervazione intorno ai follicoli piliferi: awl / auchenne (A1), zigzag (A2) e guardia (A3). I peli di Awl / Auchene sono innervati tre volte dalle terminazioni lanceolate C-LTMR (A4), A? -LTMR e A? adattamento rapido-LTMR (A6). I follicoli piliferi a zigzag sono i fusti dei capelli più corti e sono innervati sia da C-LTMR (A4) che da A? -LTMR terminazioni lanceolate (A5). I follicoli piliferi di guardia più lunghi sono innervati da A? terminazioni lanceolate longitudinali (A6) ad adattamento rapido-LTMR e sono associate ad A? adattamento lento-LTMR delle terminazioni touch dome (A7). Le proiezioni centrali di tutte queste fibre terminano in lamine distinte, ma parzialmente sovrapposte, del corno dorsale del midollo spinale (C-LTMR nella lamina II, A? -LTMR nella lamina III e A? -LTMR nella lamina IV e V). Le proiezioni di LTMR che innervano i follicoli piliferi stessi o adiacenti sono allineate per formare una colonna stretta nel corno dorsale del midollo spinale (B1 in grigio). Solo nella pelle pelosa, una sottopopolazione di terminazione libera delle fibre C innerva l'epidermide e risponde al tocco piacevole (A8). Queste fibre C-touch non rispondono al tocco nocivo e il loro percorso non è ancora noto (B2). Nella pelle glabra, il tocco innocuo è mediato da quattro tipi di LTMR. Il complesso neurite cellulare delle cellule di Merkel si trova nello strato basale dell'epidermide (C1). Questo meccanorecettore consiste in una disposizione tra molte cellule di Merkel e un terminale nervoso allargato da un singolo A? fibra. Le cellule di Merkel mostrano processi simili a dita che contattano i cheratinociti (C2). Il finale Ruffini è localizzato nel derma. È un sottile finale sensoriale incapsulato a forma di sigaro collegato ad A? fibra (C3). Il corpuscolo di Meissner connesso ad A? terminazione nervosa e si trova nelle papille dermiche. Questo meccanismo meccanico incapsulato è costituito da cellule di supporto impacchettate disposte come lamelle orizzontali circondate da tessuto connettivo (C4). Il corpuscolo paciniano è il più profondo meccanorecettore. Una sola A? La terminazione nervosa non mielinizzata termina al centro di questo grande corpuscolo ovoidale costituito da lamelle concentriche. Le proiezioni di queste fibre A? -LTMR nel midollo spinale sono divise in due rami. Il ramo centrale principale (B3) sale nel midollo spinale dorsale ipsilaterale formatura fascicoli cuneato o gracili (B5) su livello midollo dove i afferenti primari effettuano il primo sinapsi (B6). I neuroni secondari producono una decussazione sensoriale (B7) per formare un tratto sul lemnisco mediale che ascende attraverso il tronco cerebrale al mesencefalo, in particolare nel talamo. La branca secondaria di LTMR termina nel corno dorsale nella lamina II, IV, V e interferisce con la trasmissione del dolore (B4). Il contatto nocivo è rilevato dalla terminazione nervosa libera nell'epidermide di entrambi i peli (A9) e della pelle glabra (C7). Questi meccanocettori sono la fine di A? -HTMR e C-HTMR a stretto contatto con i cheratinociti vicini (C6). Un? -HTMR termina nella lamina I e V; C-HTMR termina nella lamina I e II (B8). A livello del corno dorsale del midollo spinale, gli HTMR primari afferenti formano sinapsi con neuroni secondari che attraversano la linea mediana e si arrampicano verso la struttura cerebrale superiore nel fascio anterolaterale (B9, B10).
Recentemente, Ginty e collaboratori hanno utilizzato una combinazione di etichettatura molecolare-genetica e approcci di tracciamento retrogrado somatotopico per visualizzare l'organizzazione delle terminazioni assonali periferiche e centrali degli LTMR nei topi.15 Le loro scoperte supportano un modello in cui le caratteristiche individuali di uno stimolo tattile complesso sono estratto dai tre tipi di follicolo pilifero e convogliato attraverso le attività di combinazioni uniche di fibre A? -, A? - e C- al corno dorsale.
Hanno dimostrato che l'etichettatura genetica dei neuroni DRG tirosina idrossilasi positivi (TH +) caratterizza una popolazione di neuroni sensoriali non peptidergici di piccolo diametro e consente la visualizzazione delle terminazioni periferiche C-LTMR nella pelle. Sorprendentemente, è stato scoperto che i rami assoneali dei singoli C-LTMR arborizzano e formano terminazioni lanceolate longitudinali intimamente associate a zigzag (80% delle terminazioni) e punteruolo / auchene (20% delle terminazioni), ma non follicoli piliferi tylotrich [Fig. 1 (A4)]. A lungo si è pensato che le terminazioni lanceolate longitudinali appartenessero esclusivamente alle A? -LTMR e quindi era inaspettato che le terminazioni delle C-LTMR formassero terminazioni lanceolate longitudinali.15 Queste C-LTMR hanno un adattamento intermedio rispetto all'adattamento lento e rapido meccanocettori mielinizzati [Fig. 2 (C1)].
Immagine 2. Recettori tattili nei mammiferi: i recettori tattili cutanei si differenziano in un tocco innocuo supportato da più recettori con bassa soglia meccanica (LTMR) nella pelle glabra e pelosa e tocco nocivo supportato da un recettore ad alta soglia meccanica (HTMR). Costituiscono terminazioni senza nervi che terminano principalmente nell'epidermide. (A) Pelle glabra. A1: I corpuscoli di Meissner rilevano il movimento della pelle e lo scivolamento dell'oggetto nella mano. Sono importanti per maneggiare oggetti e destrezza. I recettori si adattano rapidamente allo stimolo, sono collegati ad A? fibre e scarsamente a fibre C e hanno un ampio campo recettoriale. A2: I corpuscoli Ruffini rilevano l'allungamento della pelle e sono importanti per rilevare la posizione delle dita e la mano dell'oggetto. Il recettore si adatta lentamente allo stimolo e mantiene l'attività fintanto che lo stimolo è stato applicato. I recettori sono collegati ad A? fibre e hanno un ampio campo recettivo. A3: I corpuscoli paciniani sono più profondi nel derma e rilevano le vibrazioni. I recettori sono collegati ad A? fibre; si adattano rapidamente agli stimoli e hanno il campo ricettivo più ampio. (B) Tutta la pelle. B1: I complessi delle cellule di Merkel sono presenti sia nella pelle glabra che intorno ai capelli. Sono densamente espressi nella mano e sono importanti per la percezione della consistenza e la migliore discriminazione tra due punti. Sono responsabili della precisione delle dita. I recettori sono collegati ad A? fibre; si adattano lentamente agli stimoli e hanno un breve campo recettivo. B2: HTMR tattili nocivi con adattamento molto lento allo stimolo, cioè attivi fintanto che viene applicato lo stimolo nocicettivo. Sono formati dalla terminazione nervosa libera di A? e fibre C associate ai cheratinociti. (C) Pelle pelosa. C1: I follicoli piliferi sono associati ai diversi tipi di capelli. Nei topi i peli di Guard sono i più lunghi e scarsamente espressi, il punteruolo / auchenne sono di media grandezza e lo zigzag è il pelo più piccolo e più densamente espresso. Sono collegati ad A? fibre ma anche ad A? e fibre C-LTMR per capelli awl / auchenne e zizag. Rilevano il movimento dei capelli compreso il tocco piacevole durante la carezza. Si adattano rapidamente o con cinetica intermedia allo stimolo. C2: le terminazioni nervose C-touch corrispondono a un sottotipo di estremità delle fibre C con estremità libera caratterizzata da una bassa soglia meccanica. Dovrebbero codificare per sensazioni piacevoli indotte dalla carezza. Si adattano moderatamente agli stimoli e hanno un breve campo recettivo. I presunti canali ionici meccanosensibili (MS) espressi nei diversi recettori tattili sono indicati in base ai dati preliminari e riassumono le attuali ipotesi in corso di valutazione.
Una seconda grande popolazione identificata riguarda le terminazioni A? -LTMR nei follicoli Awl / Auchenne e a zigzag da confrontare con il follicolo pilifero Down ampiamente studiato nel gatto e nel coniglio. Ginty e collaboratori hanno dimostrato che TrkB è espresso a livelli elevati in un sottoinsieme di neuroni DRG di medio diametro. Le registrazioni intracellulari utilizzando la preparazione del nervo cutaneo ex vivo di fibre etichettate hanno rivelato che esse esibiscono le proprietà fisiologiche delle fibre precedentemente studiate nel gatto e nel coniglio: squisita sensibilità meccanica (soglia di Von Frey <0.07 mN), adattamento rapido delle risposte agli stimoli soprasoglia, conduzione intermedia velocità (5.8 0.9 m / s) e punte di soma strette e non piegate.15 Questi A? -LTMR formano terminazioni lanceolate longitudinali associate praticamente a ogni follicolo pilifero a zig-zag e punteruolo / auchene del tronco [Fig. 1 (A5)].
Infine, hanno mostrato che le terminazioni periferiche dell'adattamento rapido di A? Gli LTMR formano terminazioni lanceolate longitudinali associate a follicoli piliferi di guardia (o tylotrich) e punteruolo / auchene [Fig. 1 (A6)] 15 Inoltre, i peli di guardia sono anche associati a un complesso di cellule Merkel che formano una cupola tattile collegata ad A? adattando lentamente l'LTMR [Fig. 1 (A7)].
In sintesi, praticamente tutti i follicoli piliferi a zigzag sono innervati sia dalle terminazioni lanceolate C-LTMR che A? -LTMR; i peli di punteruolo / auchene sono innervati tre volte da A? terminazioni lanceolate ad adattamento rapido-LTMR, A? -LTMR e C-LTMR; I follicoli piliferi di guardia sono innervati da A? adattando rapidamente le terminazioni lanceolate longitudinali LTMR e interagiscono con A? adattamento lento-LTMR delle terminazioni touch dome. Pertanto, ogni follicolo pilifero di topo riceve combinazioni uniche e invarianti di terminazioni LTMR corrispondenti a organi terminali meccanosensoriali neurofisiologicamente distinti. Considerando la disposizione iterativa di questi tre tipi di capelli, Ginty e collaboratori propongono che la pelle pelosa consiste nella ripetizione iterativa dell'unità periferica contenente, (1) uno o due peli di guardia posizionati centralmente, (2) ~ 20 peli di punteruolo / auchenne circostanti e (3 ) ~ 80 peli a zigzag intervallati [Fig. 2 (C1)].
Proiezione del midollo spinale. Le proiezioni centrali di A? LTMR ad adattamento rapido, A? -LTMR e C-LTMR terminano in lamine distinte, ma parzialmente sovrapposte (II, III, IV) del corno dorsale del midollo spinale. Inoltre, i terminali centrali degli LTMR che innervano gli stessi follicoli piliferi o adiacenti all'interno di un'unità LTMR periferica sono allineati per formare una stretta colonna LTMR nel corno dorsale del midollo spinale [Fig. 1 (B1)]. Pertanto, sembra probabile che un cuneo, o colonna di terminazioni afferenti sensoriali primarie organizzate somatotopicamente nel corno dorsale, rappresenti l'allineamento delle proiezioni centrali di A? -, A? - e C-LTMR che innervano la stessa unità periferica e rilevano la meccanica stimoli che agiscono sullo stesso piccolo gruppo di follicoli piliferi. Sulla base del numero di peli di guardia, punteruolo / auchene e zigzag del tronco e degli arti e dei numeri di ciascun sottotipo LTMR, Ginty e collaboratori stimano che il corno dorsale del topo contenga 2,000 colonne LTMR, che corrisponde al numero approssimativo di colonne LTMR Unità LTMR 4,000
Inoltre, gli assoni dei sottotipi LTMR sono strettamente associati tra loro, avendo proiezioni intrecciate e terminazioni lanceolate interdigitate che innervano lo stesso follicolo pilifero. Inoltre, poiché i tre tipi di follicolo pilifero presentano forme, dimensioni e composizioni cellulari diverse, è probabile che abbiano distinte proprietà di regolazione della deflessione o vibrazionale. Questi risultati sono coerenti con le classiche misurazioni neurofisiologiche nel gatto e nel coniglio che indicano che A? Gli RA-LTMR e gli A? -LTMR possono essere attivati in modo differenziale dalla deflessione di diversi tipi di follicolo pilifero.16,17
In conclusione, il tatto nella pelle pelosa è la combinazione di: (1) i numeri relativi, le distribuzioni spaziali uniche e le proprietà morfologiche e deflesse distinte dei tre tipi di follicoli piliferi; (2) le combinazioni uniche di terminazioni del sottotipo LTMR associate a ciascuno dei tre tipi di follicolo pilifero; e (3) sensibilità distinte, velocità di conduzione, pattern del treno di picco e proprietà di adattamento delle quattro principali classi di LTMR associati al follicolo pilifero che consentono al sistema meccanosensoriale della pelle pelosa di estrarre e trasmettere al SNC le complesse combinazioni di qualità che definiscono un toccare.
LTMR Free-Nerve Endings
Generalmente, le terminazioni libere delle fibre C nella pelle sono HTMR, ma una sottopopolazione di fibre C non risponde al tocco nocivo. Questo sottoinsieme di afferenze tattili della fibra C (CT) rappresenta un tipo distinto di unità meccanorecettive non mielinizzate a bassa soglia esistenti nella pelle pelosa ma non glabra di umani e mammiferi [Fig. 1 (A8)]. 18,19 I TC sono generalmente associati alla percezione di una piacevole stimolazione tattile a contatto con il corpo.20,21
Le afferenze TC rispondono a forze di indentazione nell'intervallo 0.3-2.5 mN e sono quindi sensibili alla deformazione della pelle quanto molte delle A? afferenti.19 Le caratteristiche di adattamento delle afferenze TC sono quindi intermedie rispetto ai meccanocettori mielinizzati che si adattano lentamente e rapidamente. I campi recettivi delle afferenze TC umane sono approssimativamente di forma rotonda o ovale. Il campo è costituito da uno a nove piccoli punti reattivi distribuiti su un'area fino a 35 mm2.22 I recettori omologhi del topo sono organizzati in uno schema di macchie discontinue che coprono circa il 50-60% dell'area della pelle pelosa [Fig. 2 (C2)]. 23
L'evidenza di pazienti privi di afferenze tattili mielinizzate indica che la segnalazione nelle fibre CT attiva la corteccia insulare. Poiché questo sistema è povero nella codifica degli aspetti discriminanti del tatto, ma è adatto alla codifica del tocco lento e gentile, le fibre CT nella pelle pelosa possono far parte di un sistema per elaborare aspetti piacevoli e socialmente rilevanti del tatto. L'attivazione della fibra X24 CT può anche ha un ruolo nell'inibizione del dolore e recentemente è stato proposto che l'infiammazione o il trauma possano cambiare la sensazione trasmessa dalle LTMR in fibra C dal piacevole tocco al dolore. 25,26
Quale percorso CT-afferenti viaggia non è ancora noto [Fig. 1 (B2)], ma sono stati documentati input tattili a bassa soglia per le cellule di proiezione spinotalamiche, 27 ha prestato credibilità a segnalazioni di deficit sottili e controlaterali di rilevamento del contatto in pazienti umani a seguito della distruzione di questi percorsi dopo le procedure di cordotomia.28
LTMR in Glabrous Skin
Complessi neurite cellula di Merkel e cupola tattile. Merkel (1875) è stata la prima a fornire una descrizione istologica di ammassi di cellule epidermiche con grandi nuclei lobulati, in contatto con presunte fibre nervose afferenti. Presumeva che subissero il senso del tatto chiamandoli Tastzellen (cellule tattili). Negli esseri umani, i complessi di cellule di Merkel e neurite sono arricchiti nelle aree sensibili al tatto della pelle, si trovano nello strato basale dell'epidermide nelle dita, nelle labbra e nei genitali. Esistono anche nella pelle pelosa a densità inferiore. Il complesso delle cellule di Merkel è costituito da una cellula di Merkel in stretta apposizione a un terminale nervoso allargato da un singolo A mielinizzato? fibra [Fig. 1 (C1)] (recensione in Halata e collaboratori) .29 Sul lato epidermico la cellula di Merkel mostra processi simili a dita che si estendono tra i cheratinociti vicini [Fig. 1 (C2)]. Le cellule di Merkel sono cellule epidermiche derivate dai cheratinociti.30,31 Il termine touch dome è stato introdotto per nominare la grande concentrazione di complessi cellulari di Merkel nella pelle pelosa della zampa anteriore del gatto. Una cupola tattile potrebbe avere fino a 150 cellule Merkel innervate da una singola fibra A? E negli esseri umani oltre alle fibre A?, A? e anche le fibre C erano regolarmente presenti. 32-34
La stimolazione dei complessi neuritici delle cellule di Merkel si traduce in risposte di tipo I (SA I) che si adattano lentamente, che provengono da campi recettivi punteggiati con bordi taglienti. Non c'è scarica spontanea. Questi complessi rispondono alla profondità di indentazione della pelle e hanno la più alta risoluzione spaziale (0.5 mm) dei meccanocettori cutanei. Trasmettono un'immagine spaziale precisa degli stimoli tattili e si propone che siano responsabili della discriminazione di forma e tessitura [Fig. 2 (B1)]. I topi privi di cellule Merkel non possono rilevare le superfici ruvide con i piedi mentre lo fanno usando i baffi
Se la cellula di Merkel, il neurone sensoriale o entrambi siano siti di meccanotrasduzione è ancora oggetto di dibattito. Nei ratti, la distruzione fototossica delle cellule Merkel abolisce la risposta SA I.36 Nei topi con cellule Merkel geneticamente soppresse, la risposta SA I registrata nella preparazione ex vivo della pelle / nervo è completamente scomparsa, dimostrando che le cellule Merkel sono necessarie per la corretta codifica della Merkel risposte dei recettori.37 Tuttavia, la stimolazione meccanica delle cellule di Merkel isolate in coltura mediante pressione motrice non genera correnti meccanicamente controllate.38,39 I cheratinociti possono svolgere un ruolo importante nel normale funzionamento del complesso delle cellule di Merkel e neurite. I processi simili a dita delle cellule di Merkel possono muoversi con la deformazione della pelle e il movimento delle cellule dell'epidermide, e questo potrebbe essere il primo passo della trasduzione meccanica. Chiaramente, le condizioni necessarie per studiare la sensibilità meccanica delle cellule Merkel devono ancora essere stabilite.
Finali Ruffini. Le terminazioni Ruffini sono sottili terminazioni sensoriali incapsulate a forma di sigaro collegate ad A? terminazioni nervose. Le terminazioni Ruffini sono piccoli cilindri di tessuto connettivo disposti lungo filamenti di collagene dermico che sono forniti da una a tre fibre nervose mielinizzate di 4-6 m di diametro. Fino a tre cilindri di diverso orientamento nel derma possono fondersi per formare un recettore [Fig. 1 (C3)]. Strutturalmente, le terminazioni Ruffini sono simili agli organi del tendine del Golgi. Sono ampiamente espressi nel derma e sono stati identificati come meccanocettori cutanei di tipo II (SA II) ad adattamento lento. Sullo sfondo dell'attività nervosa spontanea, una scarica regolare che si adatta lentamente è suscitata da una stimolazione meccanica perpendicolare mantenuta a bassa forza o più efficacemente dall'allungamento del derma. La risposta SA II proviene da ampi campi recettivi con confini oscuri. I recettori Ruffini contribuiscono alla percezione della direzione del movimento dell'oggetto attraverso lo schema di stiramento della pelle [Fig. 2 (A2)].
Nei topi, le risposte SA I e SA II possono essere separate elettrofisiologicamente nella preparazione della pelle nervosa ex-vivo. 40 Nandasena e collaboratori hanno segnalato l'immunolocalizzazione di acquaporina 1 (AQP1) nelle terminazioni Ruffini parodontali degli incisivi di ratto suggerendo che AQP1 è coinvolto in il mantenimento dell'equilibrio osmotico dentale necessario per la meccanotrasduzione.41 Anche le terminazioni parodontali Ruffini esprimevano il canale ionico meccanosensibile putativo ASIC3.42
Corpuscoli Meissner. I corpuscoli di Meissner sono localizzati nelle papille dermiche della pelle glabra, principalmente nei palmi delle mani e nelle piante dei piedi, ma anche nelle labbra, nella lingua, nel viso, nei capezzoli e nei genitali. Anatomicamente, sono costituiti da una terminazione nervosa incapsulata, la capsula essendo costituita da cellule di supporto appiattite disposte come lamelle orizzontali incorporate nel tessuto connettivo. C'è una sola fibra nervosa A? afferenze connesse per corpuscolo [Fig. 1 (C4)]. Qualsiasi deformazione fisica del corpuscolo innesca una raffica di potenziali d'azione che cessa rapidamente, ovvero si adattano rapidamente ai recettori. Quando lo stimolo viene rimosso, il corpuscolo riacquista la sua forma e nel farlo produce un'altra raffica di potenziali d'azione. A causa della loro posizione superficiale nel derma, questi corpuscoli rispondono selettivamente al movimento della pelle, al rilevamento tattile di scivolamento e vibrazioni (20 Hz). Sono sensibili alla pelle dinamica, ad esempio tra la pelle e un oggetto che viene manipolato [Fig. 40 (A2)].
Corpuscoli paciniani. I corpuscoli paciniani sono i meccanocettori più profondi della pelle e sono i meccanorecettori cutanei incapsulati più sensibili del movimento della pelle. Questi grandi corpuscoli ovoidali (1 mm di lunghezza) costituiti da lamelle concentriche di tessuto connettivo fibroso e fibroblasti rivestiti da cellule di Schwann modificate piatte sono espressi nel derma profondo.43 Al centro del corpuscolo, in una cavità piena di liquido chiamata bulbo interno , termina un singolo A? terminazione nervosa non mielinizzata afferente [Fig. 1 (C5)]. Hanno un ampio campo recettivo sulla superficie della pelle con un centro particolarmente sensibile. Lo sviluppo e la funzione di diversi tipi di meccanorecettori che si adattano rapidamente sono interrotti nei topi mutanti c-Maf. In particolare, i corpuscoli paciniani sono gravemente atrofizzati.44
I corpuscoli paciniani mostrano un adattamento molto rapido in risposta alla rientranza della pelle, la scarica nervosa II (RA II) ad adattamento rapido che sono in grado di seguire l'alta frequenza di stimoli vibratori e consentono la percezione di eventi distanti attraverso vibrazioni trasmesse.45 Corpuscolo paciniano le afferenze rispondono a una rientranza sostenuta con attività transitoria all'inizio e all'offset dello stimolo. Sono anche chiamati rivelatori di accelerazione perché possono rilevare cambiamenti nella forza dello stimolo e, se la velocità di variazione dello stimolo viene alterata (come accade nelle vibrazioni), la loro risposta diventa proporzionale a questo cambiamento. I corpuscoli paciniani rilevano grossi cambiamenti di pressione e soprattutto vibrazioni (150 300 Hz), che possono rilevare anche a centimetri di distanza [Fig. 2 (A3)].
La risposta tonica è stata osservata nel corpuscolo pacino decapsulato.46 Inoltre, i corpuscoli Paciniani intatti rispondono con attività sostenuta durante stimoli indentazione costante, senza alterare le soglie meccaniche o la frequenza di risposta quando la segnalazione mediata da GABA è bloccata tra lamelle glia e una terminazione nervosa.47 Quindi, le componenti non neuronali del corpuscolo di Pacino possono avere due ruoli nel filtrare lo stimolo meccanico e nel modulare le proprietà di risposta del neurone sensoriale.
Proiezioni del midollo spinale. Le proiezioni degli A? -LTMR nel midollo spinale sono divise in due rami. Il ramo centrale principale sale nel midollo spinale nelle colonne dorsali omolaterali fino al livello cervicale [Fig. 1 (B3)]. I rami secondari terminano nel corno dorsale nelle lamine IV e interferiscono, ad esempio, con la trasmissione del dolore. Questo può attenuare il dolore come parte del controllo del cancello [Fig. 1 (B4)]. 48
A livello cervicale, gli assoni del ramo principale si separano in due tratti: il tratto mediano comprende il fascicolo gracile che trasmette informazioni dalla metà inferiore del corpo (gambe e tronco), e il tratto esterno comprende il fascicolo cuneato che trasmette informazioni dalla metà superiore del corpo (braccia e tronco) [Fig. 1 (B5)].
Le afferenze tattili primarie realizzano la loro prima sinapsi con i neuroni del secondo ordine a livello del midollo, dove le fibre di ciascun tratto sinapsi in un nucleo con lo stesso nome: la sinapsi gracile fasciculus axones nel nucleo gracile e la sinapsi cuneate axones nel nucleo cuneate [Fig. 1 (B6)]. I neuroni che ricevono la sinapsi forniscono le afferenze secondarie e attraversano immediatamente la linea mediana per formare un tratto sul lato controlaterale del tronco cerebrale, il lemnisco mediale, che ascende attraverso il tronco cerebrale alla stazione di ripetizione successiva nel mesencefalo, in particolare nel talamo [Fig. . 1 (B7)].
Specifiche molecolari di LTMR. Recentemente sono stati in parte chiariti i meccanismi molecolari che controllano la diversificazione precoce delle LTMR. Bourane e collaboratori hanno dimostrato che le popolazioni neuronali che esprimono il recettore Ret tirosina chinasi (Ret) e il suo co-recettore GFR? 2 in topi embrionali E11 13 DRG coesprimono selettivamente il fattore di trascrizione Mafa.49,50 Questi autori dimostrano che il Mafa / Neuroni Ret / GFR? 2 destinati a diventare tre tipi specifici di LTRM alla nascita: i neuroni SA1 che innervano i complessi delle cellule di Merkel, i neuroni che si adattano rapidamente che innervano i corpuscoli di Meissner e le afferenze che si adattano rapidamente (RA I) che formano terminazioni lanceolate attorno ai follicoli piliferi. Ginty e collaboratori riferiscono anche che i neuroni DRG che esprimono Ret precoce stanno rapidamente adattando meccanorecettori da corpuscoli di Meissner, corpuscoli paciniani e terminazioni lanceolate attorno ai follicoli piliferi.51 Essi innervano zone target discrete all'interno dei nuclei gracili e cuneati, rivelando uno schema meccanosensoriale specifico per modalità proiezioni assonali dei neuroni all'interno del tronco cerebrale.
Esplorazione dei meccanocettori della pelle umana. La tecnica della microneurografia descritta da Hagbarth e Vallbo nel 1968 è stata applicata per studiare il comportamento di scarica di singole terminazioni meccanosensibili umane che forniscono muscoli, articolazioni e pelle (vedi per la rassegna Macefield, 2005) .52,53 La maggior parte della microneurografia della pelle umana studi hanno caratterizzato la fisiologia delle afferenze tattili nella pelle glabra della mano. Le registrazioni di microelettrodi dai nervi mediano e ulnare in soggetti umani hanno rivelato la sensazione al tatto generata dalle quattro classi di LTMR: le afferenze di Meissner sono particolarmente sensibili al passaggio della luce attraverso la pelle, rispondendo alle forze di taglio locali e agli scivolamenti incipienti o evidenti all'interno del campo recettivo. Le afferenze paciniane sono squisitamente sensibili ai transitori meccanici vivaci. Le afferenze rispondono vigorosamente al soffiare sul campo ricettivo. Un corpuscolo paciniano situato in una cifra di solito risponde al tocco del tavolo che sostiene il braccio. Le afferenze Merkel hanno tipicamente un'elevata sensibilità dinamica agli stimoli di indentazione applicati a un'area discreta e spesso rispondono con una scarica durante il rilascio. Sebbene le afferenze Ruffini rispondano alle forze applicate normalmente sulla pelle, una caratteristica unica delle afferenze SA II è la loro capacità di rispondere anche allo stiramento laterale della pelle. Infine, le unità dei capelli nell'avambraccio hanno ampi campi ricettivi ovoidali o irregolari composti da più punti sensibili che corrispondevano ai singoli peli (ogni fornitura afferente ~ 20 capelli).
Sensibilità meccanica dei cheratinociti
Qualsiasi stimolo meccanico sulla pelle deve essere trasmesso attraverso i cheratinociti che formano l'epidermide. Queste cellule ubiquitarie possono svolgere funzioni di segnalazione in aggiunta ai loro ruoli di supporto o protettivi. Per esempio, i cheratinociti secernono ATP, un'importante molecola di segnalazione sensoriale, in risposta a stimoli meccanici e osmotici.54,55 Il rilascio di ATP induce l'aumento del calcio intracellulare mediante la stimolazione autocrina dei recettori purinergici.55 Inoltre, vi è evidenza che l'ipotonicità attiva la Rho-chinasi via di segnalazione e la conseguente formazione di fibre da stress F-actina che suggeriscono che la deformazione meccanica dei cheratinociti possa interferire meccanicamente con le cellule vicine come le cellule di Merkel per il tocco innocuo e le terminazioni libere di C-C per un tocco nocivo [Fig. 1 (C6)]. 56,57
Touch Noxious
I meccanocettori ad alta soglia (HTMR) sono C- e A epidermici? terminazioni nervose libere. Non sono associati a strutture specializzate e si osservano in entrambe le pelli pelose [Fig. 1 (A9)] e pelle glabra [Fig. 1 (C7)]. Tuttavia, il termine di terminazione nervosa libera deve essere considerato prudentemente poiché le terminazioni nervose sono sempre in stretta apposizione con cheratinociti o cellule di Langherans o melanociti. L'analisi ultrastrutturale delle terminazioni nervose rivela la presenza di un reticolo endoplasmatico ruvido, mitocondri abbondanti e vescicole a nucleo denso. Le membrane adiacenti delle cellule epidermiche sono ispessite e assomigliano alla membrana post-sinaptica nei tessuti nervosi. Si noti che le interazioni tra le terminazioni nervose e le cellule epidermiche possono essere bidirezionali poiché le cellule epidermiche possono rilasciare mediatori come ATP, interleuchina (IL6, IL10) e bradichinina e viceversa le terminazioni nervose peptidergiche possono rilasciare peptidi come CGRP o sostanza P che agiscono sulle cellule epidermiche. Gli HTMR comprendono meccano-nocicettori eccitati solo da stimoli meccanici nocivi e nocicettori polimodali che rispondono anche al calore nocivo e alla chimica esogena [Fig. 2 (B2)]. 58
Le fibre afferenti HTMR terminano sui neuroni di proiezione nel corno dorsale del midollo spinale. Gli A-HTMR contattano i neuroni del secondo ordine prevalentemente nella lamina I e V, mentre i C-HTMR terminano nella lamina II [Fig. 1 (B8)]. I neuroni nocicettivi del secondo ordine si proiettano sul lato controlaterale del midollo spinale e ascendono nella sostanza bianca, formando il sistema anterolaterale. Questi neuroni terminano principalmente nel talamo [Fig. 1 (B9 e B10)].
Mechano-Currents in Neuroni Somatosensoriali
I meccanismi di adattamento lento o rapido dei meccanocettori non sono ancora stati chiariti. Non è chiaro fino a che punto l'adattamento dei meccanorecettori sia fornito dall'ambiente cellulare della terminazione del nervo sensoriale, dalle proprietà intrinseche dei canali meccanicamente controllati e dalle proprietà dei canali ionici voltaggio-dipendenti assonali nei neuroni sensoriali (Fig. 2). Tuttavia, i recenti progressi nella caratterizzazione di correnti con gate meccanici hanno dimostrato che esistono diverse classi di canali meccanosensibili nei neuroni DRG e possono spiegare alcuni aspetti dell'adattamento dei meccanocettori.
La registrazione in vitro nei roditori ha dimostrato che il soma dei neuroni DRG è intrinsecamente meccanosensibile ed esprime correnti cationiche meccanicamente controllate.59-64 Gadolinio e rosso rutenio bloccano completamente le correnti meccanosensibili, mentre calcio e magnesio esterni, a concentrazioni fisiologiche, così come amiloride e benzamil, causano il blocco parziale.60,62,63 FM1-43 agisce come un bloccante duraturo e l'iniezione di FM1-43 nella zampa posteriore dei topi diminuisce la sensibilità al dolore nel test di Randall Selitto e aumenta la soglia di ritiro della zampa valutata con i capelli di von Frey. 65
In risposta alla stimolazione meccanica prolungata, le correnti meccanosensibili diminuiscono attraverso la chiusura. Sulla base delle costanti di tempo del decadimento della corrente, sono stati distinti quattro tipi distinti di correnti meccanosensibili: correnti di adattamento rapido (~ 3 6 ms), correnti di adattamento intermedio (~ 15 30 ms), correnti di adattamento lento (~ 200 300 ms) ) e correnti di adattamento ultra-lento (~ 1000 ms) .64 Tutte queste correnti sono presenti con incidenza variabile nei neuroni DRG di ratto che innervano la pelle glabra della zampa posteriore.64
La sensibilità meccanica delle correnti meccanosensibili può essere determinata applicando una serie di stimoli meccanici incrementali, consentendo un'analisi stimolo-corrente relativamente dettagliata.66 La relazione stimolo-corrente è tipicamente sigmoidale e l'ampiezza massima della corrente è determinata dal numero di canali che sono simultaneamente aperti.64,67 È interessante notare che la corrente meccanosensibile che si adatta rapidamente mostra una bassa soglia meccanica e un punto medio di mezza attivazione rispetto alla corrente meccanosensibile che si adatta ultra-lentamente.63,65
I neuroni sensoriali con fenotipi non nocicettivi esprimono preferenzialmente correnti meccanosensibili che si adattano rapidamente con una soglia meccanica inferiore.60,61,63,64,68 Viceversa, lentamente e ultra-lentamente si adattano le correnti meccanosensitive adattandosi a cellule putative non nocicettive.64,68 Questo suggerimento suggerito che queste correnti potrebbero contribuire alla diverse soglie meccaniche osservate in LTMR e HTMR in vivo. Sebbene questi esperimenti in vitro vadano presi con cautela, il supporto per la presenza nel soma dei neuroni DRG dei trasduttori meccanici a bassa e alta soglia è stato fornito anche dalla stimolazione a base di stiramento radiale di neuroni sensoriali di topo coltivati.69 Questo paradigma ha rivelato due principali popolazioni di neuroni sensibili allo stiramento, uno che risponde all'ampiezza dello stimolo basso e un altro che risponde selettivamente all'ampiezza dello stimolo elevato.
Questi risultati hanno implicazioni meccanicistiche importanti, ma speculative: la soglia meccanica dei neuroni sensoriali potrebbe avere poco a che fare con l'organizzazione cellulare del meccanorecettore ma potrebbe trovarsi nelle proprietà dei canali ionici meccanicamente controllati.
I meccanismi che sono alla base della desensibilizzazione delle correnti cationiche meccanosensibili nei neuroni DRG di ratto sono stati recentemente svelati.64,67 Risulta da due meccanismi concorrenti che influenzano le proprietà del canale: adattamento e inattivazione. L'adattamento è stato segnalato per la prima volta negli studi sulle cellule ciliate uditive. Può essere descritto operativamente come una semplice traslazione della curva di attivazione del canale del trasduttore lungo l'asse dello stimolo meccanico.70-72 L'adattamento consente ai recettori sensoriali di mantenere la loro sensibilità a nuovi stimoli in presenza di uno stimolo esistente. Tuttavia, una frazione sostanziale delle correnti meccanosensibili nei neuroni DRG non può essere riattivata a seguito di stimolazione meccanica condizionata, indicando l'inattivazione di alcuni canali trasduttori.64,67 Pertanto, sia l'inattivazione che l'adattamento agiscono in tandem per regolare le correnti meccanosensibili. Questi due meccanismi sono comuni a tutte le correnti meccanosensibili identificate nei neuroni DRG di ratto, suggerendo che gli elementi fisico-chimici correlati determinano la cinetica di questi canali.64
In conclusione, determinare le proprietà delle correnti meccanosensibili endogene in vitro è cruciale nella ricerca di identificare i meccanismi di trasduzione a livello molecolare. La variabilità osservata nella soglia meccanica e la cinetica di adattamento delle diverse correnti meccanicamente controllate nei neuroni DRG suggeriscono che le proprietà intrinseche dei canali ionici possono spiegare, almeno in parte, la soglia meccanica e la cinetica di adattamento dei meccanocettori descritti nei decenni 1960 80 utilizzando preparati ex vivo.
Proteine putensibili del meccanismo
Le correnti di ioni meccanosensibili nei neuroni somatosensoriali sono ben caratterizzate, al contrario, poco si sa sull'identità delle molecole che mediano la meccanotrasduzione nei mammiferi. Gli schermi genetici in Drosophila e C. elegans hanno identificato molecole candidate di meccanotrasduzione, tra cui le famiglie TRP e degenerina / epiteliale Na + canale (Deg / ENaC) .73 I recenti tentativi di chiarire le basi molecolari della meccanotrasduzione nei mammiferi si sono in gran parte focalizzati sugli omologhi di questi candidati . Inoltre, molti di questi candidati sono presenti nei meccanocettori cutanei e nei neuroni somatosensoriali (Fig. 2).
Canali ionici a rilevamento acido
Gli ASIC appartengono a un sottogruppo proton-gated della famiglia dei canali del Na + epiteliali degeneri.74 Tre membri della famiglia ASIC (ASIC1, ASIC2 e ASIC3) sono espressi in meccanocettori e nocicettori. Il ruolo dei canali ASIC è stato studiato in studi comportamentali utilizzando topi con delezione mirata dei geni del canale ASIC. La delezione di ASIC1 non altera la funzione dei meccanocettori cutanei ma aumenta la sensibilità meccanica delle afferenze che innervano l'intestino.75 I topi knockout per ASIC2 mostrano una ridotta sensibilità degli LTMR cutanei che si adattano rapidamente.76 Tuttavia, studi successivi hanno riportato una mancanza di effetti dell'eliminazione di ASIC2 su sia la meccano-nocicezione viscerale che la meccanosensazione cutanea.77 L'interruzione dell'ASIC3 diminuisce la sensibilità meccanica delle afferenze viscerali e riduce le risposte degli HTMR cutanei a stimoli nocivi.76
Il canale dei transitori
La superfamiglia TRP è suddivisa in sei sottofamiglie nei mammiferi.78 Quasi tutte le sottofamiglie TRP hanno membri legati alla meccanosensazione in una varietà di sistemi cellulari.79 Nei neuroni sensoriali dei mammiferi, tuttavia, i canali TRP sono meglio conosciuti per il rilevamento di informazioni termiche e la mediazione dell'infiammazione neurogena, e solo due canali TRP, TRPV4 e TRPA1, sono stati implicati nella reattività del tocco. Interrompere l'espressione di TRPV4 nei topi ha solo effetti modesti sulle soglie meccosensoriali acute, ma riduce fortemente la sensibilità a stimoli meccanici nocivi.80,81 TRPV4 è un determinante cruciale nel modellare la risposta dei neuroni nocicettivi allo stress osmotico e all'iperalgesia meccanica durante l'infiammazione.82,83 TRPA1 sembra avere un ruolo nell'iperalgesia meccanica. Topi con deficit di TRPA1 presentano ipersensibilità al dolore. TRPA1 contribuisce alla trasduzione di stimoli meccanici, freddi e chimici nei neuroni sensitivi nocicettori, ma sembra che non sia essenziale per la trasduzione delle cellule ciliate.84,85
Non vi è alcuna prova chiara che indichi che i canali TRP e i canali ASIC espressi nei mammiferi siano meccanicamente controllati. Nessuno di questi canali espressi eterologamente ricapitola la firma elettrica delle correnti meccanosensitive osservate nel loro ambiente nativo. Ciò non esclude la possibilità che i canali ASIC e TRP siano meccanotrasduttori, data l'incertezza se un canale di meccanotrasduzione possa funzionare al di fuori del suo contesto cellulare (vedere la sezione su SLP3).
Proteine piezo
Piezo protiens è stato recentemente identificato come promettente candidato per proteine di meccanosensing da Coste e collaboratori.86,87 I vertebrati hanno due membri Piezo, Piezo 1 e Piezo 2, precedentemente noti come FAM38A e FAM38B, che sono ben conservati negli eucarioti multi cellulari. Il Piezo 2 è abbondante nei DRG, mentre il Piezo 1 è appena rilevabile. Le correnti meccanosensibili piezoelettriche sono inibite da gadolinio, rosso rutenio e GsMTx4 (una tossina dalla tarantola Grammostola spatulata) .88 L'espressione di Piezo 1 o Piezo 2 in sistemi eterologhi produce correnti meccanosensibili, la cinetica di inattivazione della corrente di Piezo 2 è più veloce di Piezo 1. Analogamente alle correnti meccanosensibili endogene, le correnti piezoelettriche hanno potenziali di inversione attorno a 0 mV e non sono cationiche selettive, con Na +, K +, Ca2 + e Mg2 + tutte che permeano il canale sottostante. Allo stesso modo, le correnti piezo-dipendenti sono regolate dal potenziale di membrana, con un marcato rallentamento della cinetica corrente a potenziale depolarizzato.86
Le proteine piezoelettriche sono indubbiamente proteine di meccanosensing e condividono molte proprietà di correnti meccanosensibili che si adattano rapidamente nei neuroni sensoriali. Il trattamento dei neuroni DRG coltivati con RNA interferente corto Piezo 2 ha ridotto la proporzione di neuroni con corrente rapidamente adattabile e diminuito la percentuale di neuroni meccanosensibili.86 I domini transmembrana si trovano in tutte le proteine piezo ma non sono stati osservati evidenti motivi contenenti poro o tracce di canali ionici identificato. Tuttavia, la proteina Piezo 1 del mouse purificata e ricostituita in doppio strato lipidico asimmetrico e liposoma forma canali ionici sensibili al rosso del rutenio. 87 Un passo essenziale per convalidare la meccanotrasduzione attraverso i canali piezoelettrici è utilizzare approcci in vivo per determinare l'importanza funzionale nella segnalazione del tocco. Le informazioni sono state fornite in Drosophila dove la delezione del singolo membro Piezo ha ridotto la risposta meccanica a stimoli nocivi, senza influenzare il normale tocco. 89 Sebbene la loro struttura resti da determinare, questa nuova famiglia di proteine meccanosensitive è un soggetto promettente per la ricerca futura, oltre il confine di sensazione tattile. Ad esempio, un recente studio su pazienti con anemia (xerocitosi ereditaria) mostra il ruolo di Piezo 1 nel mantenimento dell'omeostasi del volume degli eritrociti.90
Transmembrane Channel-Like (TMC)
Uno studio recente indica che due proteine, TMC1 e TMC2, sono necessarie per la meccanotrasduzione delle cellule dei capelli. 91 La sordità ereditaria dovuta alla mutazione del gene TMC1 è stata riportata in umani e topi. 92,93 La presenza di questi canali non era ancora stata dimostrata nel sistema somatosensoriale, ma sembra essere un buon modo per indagare.
Proteina simile alla Stomatina 3 (SLP3)
Oltre ai canali di trasduzione, è stato dimostrato che alcune proteine accessorie collegate al canale hanno un ruolo nella sensibilità al tatto. SLP3 è espresso nei neuroni DRG dei mammiferi. Studi con topi mutanti privi di SLP3 hanno mostrato cambiamenti nelle correnti di meccanosensazione e meccanosentive. La funzione precisa di 94,95 SLP3 rimane sconosciuta. Può essere un linker tra il canale mechanosensitive e i microtubuli sottostanti, come proposto per il suo omologo C. elegans MEC2.96 Recentemente GR. Il laboratorio di Lewin ha suggerito che un legatore è sintetizzato dai neuroni sensoriali del DRG e collega il canale ionosferico meccanosensibile alla matrice extracellulare.97 Interrompendo il collegamento si abolisce la corrente meccanosensibile RA che suggerisce che alcuni canali ionici sono meccanosensibili solo quando legati. Le correnti RA-meccanosensitive sono inoltre inibite dalla laminina-332, una proteina matrice prodotta dai cheratinociti, rinforzando l'ipotesi di una modulazione della corrente meccanosensibile da parte di proteine extracellulari.98
Sottofamiglia canale K +
In parallelo alle correnti meccanosensibili depolarizzanti cationiche, la presenza di correnti K + riposte meccanosensitive è sotto esame. I canali K + nelle cellule meccanosensibili possono entrare nel bilancio attuale e contribuire a definire la soglia meccanica e il decorso temporale dell'adattamento dei meccanocettori.
I membri di KCNK appartengono alla famiglia del canale K + a due pori (K2P). 99,100 Il K2P mostra un notevole intervallo di regolazione da parte di agenti cellulari, fisici e farmacologici, compresi i cambiamenti di pH, il calore, l'allungamento e la deformazione della membrana. Questi K2P sono attivi a potenziale di membrana a riposo. Diverse subunità KCNK sono espresse in neuroni somatosensoriali. 101 KCNK2 (TREK-1), KCNK4 (TRAAK) e TREK-2 sono tra i pochi canali per i quali è stato dimostrato un gating meccanico diretto mediante stiramento della membrana. 102,103
I topi con un gene KCNK2 interrotto hanno mostrato una maggiore sensibilità al calore e lievi stimoli meccanici, ma una normale soglia di ritiro alla pressione meccanica nociva applicata alla zampa posteriore utilizzando il test di Randall Selitto.104 I topi carenti di KCNK2 mostrano anche un'iperalgesia termica e meccanica aumentata nell'infiammazione condizioni. I topi knockout KCNK4 erano ipersensibili alla stimolazione meccanica lieve e questa ipersensibilità è stata aumentata dall'ulteriore inattivazione di KCNK2.105 L'aumento della meccanosensibilità di questi topi knockout potrebbe significare che lo stiramento normalmente attiva correnti meccanosensibili sia depolarizzanti che ripolarizzanti in modo coordinato, analogamente allo squilibrio di depolarizzazione e ripolarizzazione delle correnti sotto tensione.
KCNK18 (TRESK) è un importante contributo alla conduttanza K + di fondo che regola il potenziale di membrana a riposo dei neuroni somatosensoriali.106 Sebbene non sia noto se KCNK18 sia direttamente sensibile alla stimolazione meccanica, può svolgere un ruolo nella mediazione delle risposte al tocco leggero, così come stimoli meccanici dolorosi. Si propone che KCNK18 e, in misura minore, KCNK3, siano il bersaglio molecolare dell'idrossi -? - sanshool, un composto trovato nei grani di pepe di Schezuan che attiva i recettori tattili e induce una sensazione di formicolio negli esseri umani.107,108
Il canale K + dipendente dalla tensione KCNQ4 (Kv7.4) è cruciale per impostare la preferenza di velocità e frequenza di una sottopopolazione di meccanorecettori ad adattamento rapido sia nei topi che nell'uomo. La mutazione di KCNQ4 è stata inizialmente associata a una forma di sordità ereditaria. È interessante notare che uno studio recente localizza KCNQ4 nelle terminazioni nervose periferiche del follicolo capillare cutaneo adattandosi rapidamente e del corpuscolo di Meissner. Di conseguenza, la perdita della funzione KCNQ4 porta ad un miglioramento selettivo della sensibilità del mechanoreceptor alle vibrazioni a bassa frequenza. In particolare, le persone con ipoacusia a insorgenza tardiva a causa di mutazioni dominanti del gene KCNQ4 mostrano prestazioni migliorate nella rilevazione di vibrazioni di piccola ampiezza e basse frequenze.109
Insight di Dr. Alex Jimenez
Il tatto è considerato uno dei sensi più complessi nel corpo umano, in particolare perché non esiste un organo specifico che lo gestisca. Invece, il senso del tatto si manifesta attraverso i recettori sensoriali, noti come meccanorecettori, che si trovano attraverso la pelle e rispondono alla pressione o alla distorsione meccanica. Ci sono quattro tipi principali di meccanocettori nella pelle glabra, o senza pelo, dei mammiferi: corpuscoli lamellari, corpuscoli tattili, terminazioni nervose di Merkel e corpuscoli bulbosi. I meccanocettori funzionano per consentire la rilevazione del tatto, al fine di monitorare la posizione dei muscoli, delle ossa e delle articolazioni, nota come propriocezione, e persino di rilevare i suoni e il movimento del corpo. Comprendere i meccanismi di struttura e funzione di questi meccanorecettori è un elemento fondamentale nell'utilizzo di trattamenti e terapie per la gestione del dolore.
Conclusione
Il tatto è un senso complesso perché rappresenta diverse qualità tattili, ovvero vibrazione, forma, consistenza, piacere e dolore, con diverse prestazioni discriminanti. Fino ad ora, la corrispondenza tra un organo tattile e il senso psicofisico era correlativa e i marcatori molecolari specifici della classe stanno appena emergendo. Lo sviluppo di test sui roditori che corrispondono alla diversità del comportamento tattile è ora richiesto per facilitare l'identificazione futura della genomica. L'uso di topi privi di specifici sottoinsiemi di tipi afferenti sensoriali faciliterà enormemente l'identificazione dei meccanocettori e delle fibre afferenti sensoriali associate a una particolare modalità tattile. È interessante notare che un recente articolo apre l'importante questione delle basi genetiche dei tratti meccanosensoriali nell'uomo e suggerisce che la singola mutazione genica potrebbe influenzare negativamente la sensibilità al tocco. 110 Questo sottolinea che la fisiopatologia del deficit tattile umano è in gran parte sconosciuta e certamente progredire identificando con precisione il sottoinsieme dei neuroni sensoriali legati a una modalità tattile oa un deficit tattile.
In cambio, sono stati fatti progressi per definire le proprietà biofisiche delle correnti mechano-gate.64 Lo sviluppo di nuove tecniche negli ultimi anni, consentendo il monitoraggio delle variazioni di tensione della membrana, mentre si registra la corrente mechano-gated, si è dimostrato un metodo sperimentale prezioso per descrivere correnti meccanosensitive con adattamento rapido, intermedio e lento (rivisto in Delmas e collaboratori) .66,111 Il futuro sarà determinare il ruolo delle proprietà correnti nei meccanismi di adattamento di meccanocettori funzionalmente diversi e il contributo di correnti K + meccanosensitive all'eccitabilità di LTMR e HTMR.
La natura molecolare delle correnti mechano-gate nei mammiferi è anche un futuro argomento di ricerca promettente. La ricerca futura progredirà in due prospettive, in primo luogo per determinare il ruolo della molecola accessoria che legherà i canali al citoscheletro e sarebbe necessaria per conferire o regolare la meccanosensibilità dei canali ionici simili delle famiglie TRP e ASIC / EnaC. In secondo luogo, per indagare l'ampia e promettente area del contributo dei canali piezoelettrici rispondendo a domande chiave, relative ai meccanismi di permeazione e gating, il sottoinsieme di neuroni sensoriali e le modalità tattili che coinvolgono Piezo e il ruolo del Piezo in cellule non neuronali associate a mechanosensation.
Il senso del tatto, rispetto a quello della vista, del gusto, del suono e dell'olfatto, che utilizzano organi specifici per elaborare queste sensazioni, può manifestarsi in tutto il corpo attraverso minuscoli recettori noti come meccanocettori. Diversi tipi di meccanocettori possono essere trovati in vari strati della pelle, dove possono rilevare un'ampia gamma di stimoli meccanici. L'articolo sopra descrive punti salienti specifici che dimostrano il progresso dei meccanismi strutturali e funzionali dei meccanocettori associati al senso del tatto. Informazioni referenziate dal National Center for Biotechnology Information (NCBI). Lo scopo delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni della colonna vertebrale. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattarci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Dolore alla schiena
Mal di schiena è una delle cause prevalenti di disabilità e giornate perse al lavoro in tutto il mondo. Di fatto, il dolore alla schiena è stato attribuito come la seconda ragione più comune per le visite di un medico, superata solo dalle infezioni delle alte vie respiratorie. Circa il 80 percento della popolazione sperimenterà qualche tipo di dolore alla schiena almeno una volta nel corso della vita. La colonna vertebrale è una struttura complessa composta da ossa, articolazioni, legamenti e muscoli, tra gli altri tessuti molli. A causa di ciò, lesioni e / o condizioni aggravate, come dischi erniciati, può eventualmente portare a sintomi di mal di schiena. Le lesioni sportive o gli incidenti automobilistici sono spesso la causa più frequente di mal di schiena, tuttavia a volte il più semplice dei movimenti può avere risultati dolorosi. Fortunatamente, le opzioni di trattamento alternative, come la cura chiropratica, possono aiutare ad alleviare il mal di schiena attraverso l'uso di aggiustamenti spinali e manipolazioni manuali, in definitiva migliorando il sollievo dal dolore.
1. Moriwaki K, Yuge O. Caratteristiche topografiche delle anomalie ipoestetiche e iperestetiche tattili cutanee nel dolore cronico.Dolore1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Croce Ref]
2. Spessore B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Sensibilizzazione meccanica e termica dei nocicettori cutanei nei ratti con neuropatia periferica sperimentaleNeuroscienze2005;132: 193-201. doi: 10.1016 / j.neuroscience.2004.12.036. [PubMed]ï ¿½[Croce Ref]
3. Kleggetveit IP, J rum E. Disfunzione delle fibre grandi e piccole nelle lesioni dei nervi periferici con o senza dolore spontaneo.J Pain2010;11: 1305-10. doi: 10.1016 / j.jpain.2010.03.004. [PubMed]ï ¿½[Croce Ref]
4. Noback CR. Morfologia e filogenesi dei capelliAnn NY Acad Sci. 1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed]ï ¿½[Croce Ref]
5. Straile WE. Follicoli piliferi atipici nella pelle del coniglioNatura1958;181:1604�5. doi: 10.1038/1811604a0.�[PubMed]ï ¿½[Croce Ref]
6. Straile WE. La morfologia dei follicoli di tylotrich nella pelle del coniglioSono J Anat. 1961;109: 1 13. doi: 10.1002 / aja.1001090102. [PubMed]ï ¿½[Croce Ref]
7. Millard CL, Woolf CJ. Innervazione sensoriale dei peli degli arti posteriori del ratto: un'analisi microscopica leggeraJ Comp Neurol. 1988;277: 183 94. doi: 10.1002 / cne.902770203. [PubMed]ï ¿½[Croce Ref]
9. Brown AG, Iggo A. Uno studio quantitativo dei recettori cutanei e delle fibre afferenti nel gatto e nel coniglio.J Physiol. 1967;193: 707 33. [Articolo gratuito di PMC]ï ¿½[PubMed]
10. Burgess PR, Petit D, Warren RM. Tipi di recettori nella pelle pelosa di gatto forniti dalle fibre mielinizzateJ Neurophysiol. 1968;31: 833 48. [PubMed]
11. Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. Le cellule della papilla dermica Sox2-positive specificano il tipo di follicolo pilifero nell'epidermide dei mammiferiSviluppo2009;136: 2815 23. doi: 10.1242 / dev.038620. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
12. Hussein MA. Lo schema generale della disposizione dei follicoli piliferi nel ratto e nel topoJ Anat. 1971;109: 307 16. [Articolo gratuito di PMC]ï ¿½[PubMed]
13. Vielkind U, Hardy MH. Cambiamento dei modelli delle molecole di adesione cellulare durante lo sviluppo del follicolo pilifero della pelatura del topo 2. Morfogenesi dei follicoli nei capelli mutanti, tabby e lanuginosiActa Anat (Basilea) 1996;157: 183 94. doi: 10.1159 / 000147880. [PubMed]ï ¿½[Croce Ref]
14. Hardy MH, Vielkind U. Cambiamento dei modelli delle molecole di adesione cellulare durante lo sviluppo del follicolo pilifero della pelatura di topo. 1. Morfogenesi dei follicoli nei topi wild-typeActa Anat (Basilea) 1996;157: 169 82. doi: 10.1159 / 000147879. [PubMed]ï ¿½[Croce Ref]
15. Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S, et al. L'organizzazione funzionale dei neuroni meccanosensoriali cutanei a bassa sogliaCell. 2011;147: 1615 27. doi: 10.1016 / j.cell.2011.11.027.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
16. Brown AG, Iggo A. Uno studio quantitativo dei recettori cutanei e delle fibre afferenti nel gatto e nel coniglio.J Physiol. 1967;193: 707 33. [Articolo gratuito di PMC]ï ¿½[PubMed]
17. Burgess PR, Petit D, Warren RM. Tipi di recettori nella pelle pelosa di gatto forniti dalle fibre mielinizzateJ Neurophysiol. 1968;31: 833 48. [PubMed]
18. Vallbo A, Olausson H, Wessberg J, Norrsell U. Un sistema di afferenze non mielinizzate per la meccanoricezione innocua nella pelle umana.Cervello Res. 1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed]ï ¿½[Croce Ref]
19. Vallbo AB, Olausson H, Wessberg J. Le afferenze non mielinizzate costituiscono un secondo sistema che codifica gli stimoli tattili della pelle pelosa umana.J Neurophysiol. 1999;81: 2753 63. [PubMed]
22. Wessberg J, Olausson H, Fernstr m KW, Vallbo AB. Proprietà del campo ricettivo delle afferenze tattili non mielinizzate nella pelle umanaJ Neurophysiol. 2003;89: 1567 75. doi: 10.1152 / jn.00256.2002. [PubMed][Croce Ref]
23. Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Visualizzazione genetica molecolare di un raro sottoinsieme di neuroni sensoriali non mielinizzati che possono rilevare un tocco delicatoNat Neurosci. 2007;10: 946 8. doi: 10.1038 / nn1937. [PubMed]ï ¿½[Croce Ref]
24. Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G, et al. Le afferenze tattili non mielinizzate segnalano il tocco e proiettano alla corteccia insulareNat Neurosci. 2002;5: 900 4. doi: 10.1038 / nn896.[PubMed]ï ¿½[Croce Ref]
25. Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. La neurofisiologia delle afferenze tattili non mielinizzate.Neurosci Biobehav Rev. 2010;34: 185. doi: 91 / j.neubiorev.10.1016. [Recensione][PubMed]ï ¿½[Croce Ref]
26. Krmer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M, et al. Attivazione della rete dolorosa corticale mediante stimolazione tattile morbida dopo iniezione di sumatriptanDolore2007;133: 72. doi: 8 / j.pain.10.1016. [PubMed]ï ¿½[Croce Ref]
27. Applebaum AE, Beall JE, Foreman RD, Willis WD. Organizzazione e campi recettivi dei neuroni del tratto spinotalamico dei primatiJ Neurophysiol. 1975;38: 572 86. [PubMed]
28. JC bianco, dolce WH. Efficacia della cordotomia nel dolore fantasma dopo l'amputazioneAMA Arch Neurol Psychiatry1952;67: 315 22. [PubMed]
29. Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel e la sua `` cellula di Merkel '', morfologia, sviluppo e fisiologia: revisione e nuovi risultati.Anat Rec A Discov Mol Cell Evol Biol. 2003;271: 225 39. doi: 10.1002 / ar.a.10029. [PubMed]ï ¿½[Croce Ref]
30. Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Le cellule di Merkel dei mammiferi discendono dal lignaggio epidermicoDev Biol. 2009;336: 76. doi: 83 / j.ydbio.10.1016.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
31. Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N, et al. I progenitori epidermici danno origine alle cellule di Merkel durante lo sviluppo embrionale e l'omeostasi degli adultiJ Cell Biol. 2009;187: 91 100. doi: 10.1083 / jcb.200907080. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
32. Ebara S, Kumamoto K, Baumann KI, Halata Z. Le analisi tridimensionali delle cupole tattili nella pelle pelosa della zampa di gatto rivelano substrati morfologici per elaborazioni sensoriali complesse.Neurosci Res. 2008;61: 159. doi: 71 / j.neures.10.1016. [PubMed]ï ¿½[Croce Ref]
33. Guinard D, Usson Y, Guillermet C, Saxod R. Merkel complessi della pelle digitale umana: imaging tridimensionale con microscopia laser confocale e doppia immunofluorescenza.J Comp Neurol. 1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Croce Ref]
34. Reinisch CM, Tschachler E. La cupola tattile nella pelle umana è fornita da diversi tipi di fibre nervoseAnn Neurol. 2005;58: 88. doi: 95 / ana.10.1002. [PubMed]ï ¿½[Croce Ref]
35. Maricich SM, Morrison KM, Mathes EL, Brewer BM. I roditori si affidano alle cellule Merkel per i compiti di discriminazione della tramaJ Neurosci. 2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
36. Ikeda I, Yamashita Y, Ono T, Ogawa H. La distruzione fototossica selettiva delle cellule di Merkel di ratto abolisce le risposte delle unità meccanorecettrici di tipo I. che si adattano lentamente.J Physiol. 1994;479: 247 56. [Articolo gratuito di PMC][PubMed]
37. Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, et al. Le cellule Merkel sono essenziali per risposte leggereScienza2009;324: 1580 2. doi: 10.1126 / science.1172890.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
38. Diamond J, Holmes M, infermiera CA. Le sinapsi reciproche cellula-neurite di Merkel sono coinvolte nell'attivazione delle risposte tattili nella pelle della salamandra? J Physiol. 1986;376: 101 20. [Articolo gratuito di PMC][PubMed]
39. Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Correnti dipendenti dalla tensione in singole cellule di ratto Merkel isolate.J Physiol. 1992;450: 143 62. [Articolo gratuito di PMC]ï ¿½[PubMed]
40. Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. La regolarità dello sparo prolungato rivela due popolazioni di recettori tattili che si adattano lentamente nella pelle pelosa di topoJ Neurophysiol. 2010;103: 3378 88. doi: 10.1152 / jn.00810.2009. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
41. Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T.Immunolocalizzazione di acquaporina-1 nelle terminazioni meccanorecettive di Ruffini nel legamento parodontale.Cervello Res. 2007;1157: 32. doi: 40 / j.brainres.10.1016. [PubMed]ï ¿½[Croce Ref]
42. Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K, et al. Rilevazione del canale ionico 3 sensibile all'acido (ASIC3) nelle terminazioni parodontali Ruffini degli incisivi di topo.Neurosci Lett. 2011;488: 173 7. doi: 10.1016 / j.neulet.2010.11.023. [PubMed]ï ¿½[Croce Ref]
43. Johnson KO. I ruoli e le funzioni dei meccanocettori cutanei . Curr Opin Neurobiol. 2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed]ï ¿½[Croce Ref]
44. Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A, et al. Il fattore di trascrizione c-Maf controlla lo sviluppo e la funzione del recettore tattileScienza2012;335: 1373 6. doi: 10.1126 / science.1214314. [PubMed]ï ¿½[Croce Ref]
45. Mendelson M, Lowenstein WR. Meccanismi di adattamento dei recettoriScienza1964;144: 554 5. doi: 10.1126 / science.144.3618.554. [PubMed]ï ¿½[Croce Ref]
46. Loewenstein WR, Mendelson M. Componenti dell'adattamento del recettore in un corpuscolo pacinianoJ Physiol. 1965;177: 377 97. [Articolo gratuito di PMC]ï ¿½[PubMed]
47. Pawson L, Prestia LT, Mahoney GK, G l B, Cox PJ, Pack AK. L'interazione GABAergica / glutammatergico-gliale / neuronale contribuisce al rapido adattamento nei corpuscoli paciniani.J Neurosci. 2009;29:2695�705. doi: 10.1523/JNEUROSCI.5974-08.2009.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
48. Basbaum AI, Jessell TM. La percezione del dolore. In: Kandel ER, Schwartz JH, Jessell TM, eds. Principi della scienza neurale. Quarta edizione. I compagini della McGraw-Hill, 2000: 472-490.
49. Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A, et al. I sottotipi di meccanorecettori a bassa soglia esprimono selettivamente MafA e sono specificati dalla segnalazione RetNeurone. 2009;64: 857 70. doi: 10.1016 / j.neuron.2009.12.004. [PubMed]ï ¿½[Croce Ref]
50. Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. Un ruolo per la segnalazione del fattore di trascrizione Runx nella diversificazione dei neuroni sensoriali del ganglio della radice dorsale.Neurone. 2006;49: 379 93. doi: 10.1016 / j.neuron.2006.01.008. [PubMed]ï ¿½[Croce Ref]
51. Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Identificazione molecolare di meccanocettori che si adattano rapidamente e la loro dipendenza dello sviluppo dalla segnalazione retNeurone. 2009;64: 841 56. doi: 10.1016 / j.neuron.2009.11.003. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
52. Vallbo AB, Hagbarth KE. Attività dei meccanocettori cutanei registrati per via percutanea in soggetti umani svegliExp Neurol. 1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed]ï ¿½[Croce Ref]
53. Macefield VG. Caratteristiche fisiologiche dei meccanocettori a bassa soglia nelle articolazioni, nei muscoli e nella pelle nei soggetti umaniClin Exp Pharmacol Physiol. 2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed]ï ¿½[Croce Ref]
54. Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Le onde Ca2 + nei cheratinociti vengono trasmesse ai neuroni sensoriali: il coinvolgimento dell'ATP extracellulare e l'attivazione dei recettori P2Y2.Biochem J. 2004;380: 329. doi: 38 / BJ10.1042. [Articolo gratuito di PMC]ï ¿½[PubMed][Croce Ref]
55. La segnalazione di Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. ATP è cruciale per la risposta dei cheratinociti umani alla stimolazione meccanica da shock ipoosmotico.Exp Dermatol. 2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed]ï ¿½[Croce Ref]
56. Amano M, Fukata Y, Kaibuchi K.Regolazione e funzioni della chinasi associata a RhoExp Cell Res. 2000;261: 44. doi: 51 / excr.10.1006. [Recensione] [PubMed]ï ¿½[Croce Ref]
57. Koyama T, Oike M, Ito Y. Coinvolgimento della Rho-chinasi e della tirosina chinasi nel rilascio di ATP ipotonico indotto da stress nelle cellule endoteliali dell'aorta bovina.J Physiol. 2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
59. McCarter GC, Reichling DB, Levine JD. Trasduzione meccanica da parte dei neuroni gangliari della radice dorsale di ratto in vitroNeurosci Lett. 1999;273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[PubMed]ï ¿½[Croce Ref]
60. Drew LJ, Wood JN, Cesare P. Distinte proprietà meccanosensibili dei neuroni sensoriali sensibili e insensibili alla capsaicina.J Neurosci. 2002;22: RC228. [PubMed]
61. Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P, et al. I canali ionici sensibili all'acido ASIC2 e ASIC3 non contribuiscono alle correnti attivate meccanicamente nei neuroni sensoriali dei mammiferi.J Physiol. 2004;556: 691 710. doi: 10.1113 / jphysiol.2003.058693. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
62. McCarter GC, Levine JD. Base ionica di una corrente di meccanotrasduzione nei neuroni gangliari della radice dorsale del ratto adultoMol Pain2006;2:28. doi: 10.1186/1744-8069-2-28.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
63. Coste B, Crest M, Delmas P.Dissezione farmacologica e distribuzione di NaN / Nav1.9, correnti Ca2 + di tipo T e correnti cationiche attivate meccanicamente in diverse popolazioni di neuroni DRG.J Gen Physiol. 2007;129: 57. doi: 77 / jgp.10.1085. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
64. Hao J, Delmas P. Molteplici meccanismi di desensibilizzazione dei canali meccanotrasduttori modellano l'attivazione dei neuroni meccanosensoriali.J Neurosci. 2010;30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[PubMed]ï ¿½[Croce Ref]
65. Drew LJ, Wood JN. FM1-43 è un bloccante permanente dei canali ionici meccanosensibili nei neuroni sensoriali e inibisce le risposte comportamentali agli stimoli meccanici.Mol Pain2007;3:1. doi: 10.1186/1744-8069-3-1.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
67. Rugiero F, Drew LJ, Wood JN. Proprietà cinetiche delle correnti attivate meccanicamente nei neuroni sensoriali spinaliJ Physiol. 2010;588: 301 14. doi: 10.1113 / jphysiol.2009.182360. [Articolo gratuito di PMC]ï ¿½[PubMed][Croce Ref]
68. Hu J, Lewin GR. Correnti meccanosensibili nei neuriti dei neuroni sensoriali del topo in colturaJ Physiol. 2006;577: 815 28. doi: 10.1113 / jphysiol.2006.117648. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
69. Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D.L'allungamento radiale rivela popolazioni distinte di neuroni somatosensoriali dei mammiferi meccanosensibili.Proc Natl Acad Sci US A. 2008;105: 20015 20. doi: 10.1073 / pnas.0810801105. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
70. Crawford AC, Evans MG, Fettiplace R. Attivazione e adattamento delle correnti del trasduttore nelle cellule ciliate di tartaruga.J Physiol. 1989;419: 405 34. [Articolo gratuito di PMC]ï ¿½[PubMed]
71. Ricci AJ, Wu YC, Fettiplace R. Il tampone di calcio endogeno e il decorso temporale dell'adattamento del trasduttore nelle cellule ciliate uditive.J Neurosci. 1998;18: 8261 77. [PubMed]
72. Vollrath MA, Kwan KY, Corey DP. Le micromacchine di meccanotrasduzione nelle cellule ciliateAnnu Rev Neurosci. 2007;30: 339. doi: 65 / annurev.neuro.10.1146. [Recensione] [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
74. Waldmann R, Lazdunski MH. H (+) - canali cationici con gate: sensori di acido neuronale nella famiglia di canali ionici NaC / DEG.Curr Opin Neurobiol. 1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed]ï ¿½[Croce Ref]
75. Pagina AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, et al. Il canale ionico ASIC1 contribuisce alla funzione dei meccanorecettori viscerali ma non cutaneiGastroenterologia . 2004;127: 1739 47. doi: 10.1053 / j.gastro.2004.08.061. [PubMed]ï ¿½[Croce Ref]
76. Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, et al. Il canale cationico DRASIC contribuisce alla rilevazione del tocco cutaneo e degli stimoli acidi nei topiNeurone. 2001;32: 1071. doi: 83 / S10.1016-0896 (6273) 01-00547. [Errata corrige in: Neuron 5 Jul 2002; 18] [35] [PubMed]ï ¿½[Croce Ref]
77. Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M, et al. Il knockout del canale ASIC2 nei topi non compromette la meccanosensazione cutanea, la meccanonociczione viscerale e l'udito.J Physiol. 2004;558: 659 69. doi: 10.1113 / jphysiol.2004.066001. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
78. Damann N, Voets T, Nilius B. TRP nei nostri sensiCurr Biol. 2008;18: R880 9. doi: 10.1016 / j.cub.2008.07.063. [Recensione] [PubMed]ï ¿½[Croce Ref]
80. Liedtke W, Tobin DM, Bargmann CI, Friedman JM. Il TRPV4 dei mammiferi (VR-OAC) dirige le risposte comportamentali agli stimoli osmotici e meccanici in Caenorhabditis elegans.Proc Natl Acad Sci US A. 2003;100(Suppl 2): 14531. doi: 6 / pnas.10.1073. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
81. Suzuki M, Mizuno A, Kodaira K, Imai M. Sensazione di pressione ridotta nei topi privi di TRPV4.J Biol Chem. 2003;278: 22664 8. doi: 10.1074 / jbc.M302561200. [PubMed]ï ¿½[Croce Ref]
82. Liedtke W, Choe Y, Mart -Renom MA, Bell AM, Denis CS, Sali A, et al. Canale attivato osmoticamente correlato al recettore vanilloide (VR-OAC), un osmorecettore vertebrato candidato.Cell. 2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
83. Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. Il potenziale transitorio del recettore vanilloide 4 è essenziale nel dolore neuropatico indotto dalla chemioterapia nel rattoJ Neurosci. 2004;24: 4444 52. doi: 10.1523 / JNEUROSCI.0242-04.2004. [Errata corrige in: J Neurosci. 2004 Jun; 24] [23][PubMed]ï ¿½[Croce Ref]
84. Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, et al. TRPA1 media le azioni infiammatorie di irritanti ambientali e agenti proalgesiciCell. 2006;124: 1269 82. doi: 10.1016 / j.cell.2006.02.023. [PubMed]ï ¿½[Croce Ref]
85. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ, et al. TRPA1 contribuisce alla nocicezione fredda, meccanica e chimica ma non è essenziale per la trasduzione delle cellule ciliate.Neurone. 2006;50: 277 89. doi: 10.1016 / j.neuron.2006.03.042. [PubMed]ï ¿½[Croce Ref]
86. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 e Piezo2 sono componenti essenziali di distinti canali cationici attivati meccanicamenteScienza2010;330: 55 60. doi: 10.1126 / science.1193270. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
87. Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS, et al. Le proteine piezo sono subunità che formano i pori di canali attivati meccanicamenteNatura2012;483: 176. doi: 81 / nature10.1038.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
88. Bae C, Sachs F, Gottlieb PA. Il canale ionico meccanosensibile Piezo1 è inibito dal peptide GsMTx4. Biochimica2011;50: 6295 300. doi: 10.1021 / bi200770q. [Articolo gratuito di PMC]ï ¿½[PubMed][Croce Ref]
89. Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. Il ruolo della Drosophila Piezo nella nocicezione meccanica.Natura2012;483: 209-12. doi: 10.1038 / nature10801. [Articolo gratuito di PMC]ï ¿½[PubMed][Croce Ref]
90. Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. Le mutazioni nella proteina di meccanotrasduzione PIEZO1 sono associate a xerocitosi ereditaria.Sangue2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
91. Kawashima Y, G l oc GS, Kurima K, Labay V, Lelli A, Asai Y, et al. La meccanotrasduzione nelle cellule ciliate dell'orecchio interno del topo richiede geni simili a canali transmembranaJ Clin Invest. 2011;121: 4796 809. doi: 10.1172 / JCI60405. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
92. Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouchène J, et al. TMC1 ma non TMC2 è responsabile della compromissione dell'udito non sindromica autosomica recessiva nelle famiglie tunisine.Audiol Neurootol. 2008;13: 213 8. doi: 10.1159 / 000115430. [PubMed]ï ¿½[Croce Ref]
93. Manji SS, Miller KA, Williams LH, Dahl HH. Identificazione di tre nuovi ceppi di topi ipoacusici con mutazioni nel gene Tmc1Sono J Pathol2012;180: 1560 9. doi: 10.1016 / j.ajpath.2011.12.034.[PubMed]ï ¿½[Croce Ref]
94. Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, et al. Una proteina del dominio della stomatina essenziale per la sensazione tattile nel topoNatura2007;445: 206-9. doi: 10.1038 / nature05394. [PubMed][Croce Ref]
95. Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, et al. Meccanotrasduzione della stomatina e dei neuroni sensorialiJ Neurophysiol. 2007;98: 3802 8. doi: 10.1152 / jn.00860.2007.[PubMed]ï ¿½[Croce Ref]
96. Huang M, Gu G, Ferguson EL, Chalfie M. Una proteina simile alla stomatina necessaria per la meccanosensazione in C. elegans.Natura1995;378:292�5. doi: 10.1038/378292a0.�[PubMed]ï ¿½[Croce Ref]
97. Hu J, Chiang LY, Koch M, Lewin GR. Prova di un legame proteico coinvolto nel tocco somaticoEMBO J. 2010;29: 855 67. doi: 10.1038 / emboj.2009.398. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
98. Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L, et al. La laminina-332 coordina la meccanotrasduzione e la biforcazione del cono di crescita nei neuroni sensorialiNat Neurosci. 2011;14: 993 1000. doi: 10.1038 / nn.2873. [PubMed]ï ¿½[Croce Ref]
99. Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, et al. TWIK-1, un canale K + umano onnipresente che rettifica debolmente verso l'interno con una nuova strutturaEMBO J. 1996;15: 1004 11.[Articolo gratuito di PMC]ï ¿½[PubMed]
100. Lesage F.Farmacologia dei canali del potassio di fondo neuronaliNeurofarmacologia . 2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed]ï ¿½[Croce Ref]
101. Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE, et al. Analisi della distribuzione dei canali del potassio nel dominio dei due pori umani nei tessuti del sistema nervoso centrale e della periferiaBrain Res Mol Brain Res. 2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Croce Ref]
102. Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor E. Stimolazione meccanica o acida, due modalità interattive di attivazione del canale del potassio TREK-1.J Biol Chem. 1999;274: 26691 6. doi: 10.1074 / jbc.274.38.26691. [PubMed]ï ¿½[Croce Ref]
103. Maingret F, Fosset M, Lesage F, Lazdunski M, Honor E. TRAAK è un canale K + con controllo meccanico dei neuroni dei mammiferi.J Biol Chem. 1999;274: 1381 7. doi: 10.1074 / jbc.274.3.1381. [PubMed][Croce Ref]
104. Alloui A, Zimmermann K, Mamet J, Duprat F, No l J, Chemin J, et al. TREK-1, un canale K + coinvolto nella percezione del dolore polimodaleEMBO J. 2006;25: 2368 76. doi: 10.1038 / sj.emboj.7601116.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
105. No l J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, et al. I canali K + attivati meccanicamente TRAAK e TREK-1 controllano la percezione sia del caldo che del freddoEMBO J. 2009;28: 1308 18. doi: 10.1038 / emboj.2009.57. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
106. Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. I canali K + a due domini TRESK costituiscono una componente significativa delle correnti di potassio di fondo nei neuroni gangliari della radice dorsale murina.J Physiol. 2007;585: 867 79. doi: 10.1113 / jphysiol.2007.145649.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
107. Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR, et al. Gli agenti pungenti dei peperoni di Sichuan eccitano i neuroni sensoriali inibendo i canali del potassio a due pori.Nat Neurosci. 2008;11: 772 9. doi: 10.1038 / nn.2143. [Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
108. Lennertz RC, Tsunozaki M, Bautista DM, Stucky CL. Basi fisiologiche della parestesia formicolio evocato da idrossi-alfa-sanshoolJ Neurosci. 2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
109. Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM, et al. KCNQ4 Canali K (+) sintonizzano i meccanocettori per la normale sensazione tattile nel mouse e nell'uomoNat Neurosci. 2012;15: 138 45. doi: 10.1038 / nn.2985. [PubMed]ï ¿½[Croce Ref]
110. Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG, et al. Una base genetica per i tratti meccanosensoriali negli esseri umaniPLoS Biol. 2012;10: E1001318. doi: 10.1371 / journal.pbio.1001318.[Articolo gratuito di PMC]ï ¿½[PubMed]ï ¿½[Croce Ref]
111. Delmas P, Hao J, Rodat-Despoix L. Meccanismi molecolari della meccanotrasduzione nei neuroni sensoriali dei mammiferi.Nat Rev Neurosci. 2011;12: 139 53. doi: 10.1038 / nrn2993. [PubMed]ï ¿½[Croce Ref]
Dolore meccanorecettivo: Secondo il CDC, oltre il 50% degli adulti statunitensi (125 milioni) aveva un muscolo scheletrico disturbo del dolore nel 2012.
Più del 40% degli adulti con un disturbo del dolore muscoloscheletrico ha utilizzato un approccio di salute complementare per qualsiasi motivo nel 2012. Questo era significativamente più alto dell'uso tra le persone senza un disturbo del dolore muscoloscheletrico (24.1%). L'uso di approcci sanitari complementari per qualsiasi motivo tra le persone con dolore o problemi al collo era più del doppio dell'uso tra le persone senza questi problemi.
"Tra gli adulti con un disturbo del dolore muscoloscheletrico, l'uso di qualsiasi approccio di salute complementare era più alto tra quelli con dolore o problemi al collo (50.6%), seguito da persone con altri problemi muscolo-scheletrici (46.2%)".
L'utilizzo di approcci sanitari complementari per qualsiasi motivo tra le persone con dolore o problemi al collo era più del doppio dell'uso tra le persone senza questi problemi.
I meccanocettori sono recettori sensoriali che rispondono alla pressione o alla distorsione meccanica.
Questi includono recettori cutanei per il tatto, recettori che monitorano la lunghezza e la tensione muscolare, i recettori uditivi e vestibolari e altri.
GATE CONTROL THEORY OF DOLORE
L'input non doloroso chiude le porte a input dolorosi.
Ciò impedisce alle sensazioni del dolore di spostarsi a livelli corticali più elevati
Afferenze di piccolo diametro (dolore) impediscono l'inibizione del dolore
Afferenti di grande diametro (vibrazione) tendono ad eccitare l'inibizione del dolore.
Questa teoria afferma che le fibre non nocicettive possono interferire con i segnali delle fibre del dolore, quindi, inibendo il dolore.
Grande diametro A? le fibre non sono inocicettive (non trasmettono stimoli dolorosi) e inibiscono gli effetti dello sparo di A? e fibre C.
PERCORSO LEMNISCALE DELLA MEDIA DELLA COLONNA DORSALE
UTILIZZO DI MECCANORECETTORI PERIFERICI PER ALTERA PERCEZIONE DEL DOLORE
Come possiamo aiutare?
RIFLESSO DI RECESSO
Un neurone afferente eccitato stimola gli interneuroni eccitatori che a loro volta stimolano i motoneuroni efferenti che forniscono i bicipiti, il muscolo nel braccio che si piega (si piega) l'articolazione del gomito. La contrazione del bicipite allontana la mano dalla stufa calda.
Il neurone afferente stimola anche gli interneuroni inibitori che a loro volta inibiscono i neuroni efferenti che forniscono i tricipiti per impedirne la contrazione. Questo tipo di connessione neuronale che coinvolge la stimolazione del rifornimento nervoso di un muscolo e l'inibizione simultanea dei nervi al suo muscolo antagonista è nota come inibizione reciproca.
Il neurone afferente stimola ancora altri interneuroni che trasportano il segnale sul midollo spinale al cervello attraverso un percorso ascendente. Solo quando l'impulso raggiunge l'area sensoriale della corteccia è la persona consapevole del dolore, della sua posizione e del tipo di stimolo. Inoltre, quando l'impulso raggiunge il cervello, l'informazione può essere archiviata come memoria e la persona può pensare a quello che è successo.
TERAPIA BASATA SU RECETTORI
Regolazioni
L'attivazione dei meccanocettori articolari attraverso aggiustamenti chiropratici può modulare e `` oscurare '' la percezione cerebrale delle fibre di diametro inferiore.
La ripetizione dell'attivazione dei meccanocettori articolari può creare plasticità positiva nei percorsi afferenti.
La plasticità positiva può fermare il dolore
Vibrazione
La stimolazione vibratoria a frequenze specifiche può alterare la percezione del dolore
La ripetizione dell'attivazione dei dischi di Merkel e dei corpuscoli di Meissner può creare una plasticità positiva nei percorsi afferenti.
Ancora una volta, la plasticità positiva può arrestare il dolore
VIBRAZIONI
Questo tipo di dispositivo applica vibrazioni sinusoidali e offre un'ampiezza selezionabile continua di 0-5.2 mm a seconda della posizione del piede e una frequenza selezionabile di 5-30 Hz.
La formazione WBV sembra essere un intervento efficace, sicuro e adatto per i dipendenti che lavorano seduti con lombalgia cronica .
La stimolazione vibro-tattile motopica ha prodotto una riduzione del dolore da calore del 40% in tutti i gruppi di soggetti. La distrazione non sembrava influenzare le valutazioni sperimentali del dolore
La stimolazione vibrotattile ha reclutato efficacemente meccanismi analgesici non solo nella NC ma anche nei pazienti con dolore muscoloscheletrico cronico, inclusa la FM.
TOCCO LEGGERO
In totale, 44 volontari sani hanno sperimentato dolore da calore e stimoli TC ottimali (spazzolatura lenta) e TC non ottimali (spazzolatura rapida o vibrazione). Sono stati utilizzati tre diversi paradigmi sperimentali: applicazione simultanea di dolore da calore e stimolazione tattile (sfioramento lento o vibrazione); Spazzolatura lenta, applicata per durata e intervalli variabili, precedendo il dolore termico; Spazzolatura lenta e veloce che precede il dolore da calore
Negli esseri umani, le principali aree del cervello che ricevono informazioni C-LTMR appartengono al sistema somatosensoriale e influenzano l'elaborazione delle reti cerebrali come la corteccia insulare posteriore controlaterale o la corteccia prefrontale mediale. L'intensità del tocco TAC è codificata nella corteccia somatosensoriale primaria e secondaria (S1 controlaterale, S2 bilaterale), mentre la piacevolezza è codificata nella corteccia cingolata anteriore. Le C-LTMR attivano anche regioni coinvolte nell'elaborazione della ricompensa (putamen e corteccia orbitofrontale) e nell'elaborazione di stimoli sociali (solco temporale posteriore superiore).
TUTTO IL PERIFERICO HA UNA CONSEGUENZA CENTRALE
CASE STUDY
47-year-old male ha sofferto di CVA a sinistra nel mese di ottobre di 2017.
Non ha spostato il lato destro del suo corpo dall'incidente.
Presentato alla nostra clinica perché vuole `` tornarci. ''
PUNTI SALIENTI DELL'ESAME DI FISICA
Disartria
Percezione del dolore alterata
Difficoltà con la matematica semplice
Flaccido su RUE e RLE
PUNTI SALIENTI DELL'ESAME DI FISICA
Il paziente non ha avuto movimento fino a quando non abbiamo iniziato a testare sensazioni e riflessi:
allodinia:- Si riferisce alla sensibilizzazione centrale al dolore (aumento della risposta dei neuroni) a seguito di stimolazione normalmente non dolorosa, spesso ripetitiva.
L'allodinia può causare l'innesco di una risposta al dolore da stimoli, che normalmente non provoca dolore.
La temperatura o gli stimoli fisici possono provocare allodinia, che può sembrare una sensazione di bruciore e spesso si verifica dopo una lesione a un sito.
L'alldinia è diversa dall'iperalgesia, una reazione estrema ed esagerata a uno stimolo, che è normalmente doloroso.
INTERVENTI TERAPEUTICI
Vibrazione
Tocco leggero
agopressione
Frequenze acustiche
Regolazioni!
DOPO DUE GIORNI
Terapia basata sui recettori e sul dolore meccanorecettivo
Perché il danno localizzato o la ferita causata dal trauma portano a dolore cronico e intrattabile in alcuni pazienti? Che cosa è responsabile della traduzione delle lesioni locali con dolore acuto in una condizione di dolore cronico? Perché un po 'di dolore risponde a farmaci anti-infiammatori e / o farmaci, mentre altre forme di dolore richiedono oppiacei?
Dolore è un processo complesso che coinvolge sia il sistema nervoso periferico (PNS) che il sistema nervoso centrale (SNC). La lesione tissutale innesca il PNS, che trasmette segnali attraverso il midollo spinale nel cervello, in cui si verifica la percezione del dolore. Tuttavia, che cosa causa l'intensa esperienza del dolore per svilupparsi in un fenomeno incessante? Si può fare qualcosa per impedirlo? L'evidenza lo indica il dolore cronico deriva da una combinazione di meccanismi, come i "ricordi" neurologici del dolore precedente.
Nociception: The Simplest Pathway
Il dolore acuto o nocicettivo è caratterizzato dall'esperienza regolare di disagio che si verifica in risposta a danni o lesioni di base. È protettivo, avvisandoci di allontanarci dall'origine dell'insulto e prenderci cura del trauma. I meccanismi che creano il dolore nocicettivo includono la trasduzione, che estende la stimolazione traumatica esterna in attività elettrica nei nervi afferenti primari nocicettivi specializzati. I nervi afferenti conducono quindi le informazioni sensoriali dal PNS al CNS.
Nel SNC, i dati sul dolore sono trasmessi dai neuroni sensoriali primari nelle cellule di proiezione centrale. Dopo che le informazioni sono state trasferite in tutte quelle aree del cervello che sono responsabili della nostra percezione, accade l'esperienza sensoriale reale. Il dolore nocicettivo è una reazione relativamente semplice a uno stimolo acuto particolarmente semplice. Ma i meccanici incaricati del dolore nocicettivo non possono identificare fenomeni, come il dolore che persiste nonostante la rimozione o la guarigione della stimolazione, come nel caso del dolore degli arti fantasma.
Dolore e risposta infiammatoria
In caso di lesioni più gravi, come ferite chirurgiche, i danni ai tessuti possono stimolare una reazione infiammatoria. Tuttavia, altre condizioni, in particolare l'artrite, possono anche essere caratterizzate da continui casi di infiammazione associati a sintomi di dolore intenso. I meccanismi per questo tipo di dolore correlato al danno tissutale e una risposta infiammatoria sono diversi dal dolore nocicettivo ad avvertimento precoce.
Osservando l'incisione o il sito di altri danni o lesioni, si verifica una cascata di eventi ipereccitabili nel sistema nervoso. Questo fenomeno "wind-up" corporeo inizia dalla pelle, dove è potenziato lungo i nervi periferici e culmina con una risposta di ipersensibilità lungo il midollo spinale (corno dorsale) e il cervello. Le cellule infiammatorie circondano quindi le regioni di danno tissutale e producono anche citochine e chemochine, sostanze che hanno lo scopo di mediare il processo di guarigione e di rigenerazione dei tessuti. Ma questi agenti possono anche essere considerati irritanti e regolare le proprietà dei neuroni sensoriali primari che circondano l'area del trauma.
Pertanto, i principali fattori che scatenano il dolore infiammatorio comprendono il danno ai nocicettori ad alto limite, noto come sensibilizzazione periferica, cambiamenti e alterazioni dei neuroni nel sistema nervoso e l'amplificazione dell'eccitabilità dei neuroni all'interno del sistema nervoso centrale. Questo rappresenta la sensibilizzazione centrale ed è responsabile per l'ipersensibilità, dove le aree adiacenti a quelle del vero infortunio avvertiranno dolore come se fossero ferite. Questi tessuti possono anche reagire alla stimolazione che normalmente non crea dolore, come un tocco, indossare indumenti, una leggera pressione o addirittura lavarsi i capelli, come se fossero veramente dolorosi, definiti allodinia.
Il dolore neuropatico deriva da danni o lesioni al sistema nervoso, come la sindrome del tunnel carpale, la nevralgia post-erpetica e la neuropatia diabetica. Sebbene alcuni dei meccanismi che sembrano causare dolore neuropatico si sovrappongano a quelli responsabili del dolore infiammatorio, molti di essi sono diversi e quindi necessiteranno di un approccio diverso nei confronti della loro gestione.
Il processo di sensibilizzazione periferica e centrale viene mantenuto, almeno teoricamente e sperimentalmente, durante il neurotrasmettitore eccitatorio, il glutammato, che si ritiene venga rilasciato quando viene attivato il recettore N-metil-D-aspartato (NMDA).
Il sistema nervoso è costituito da neurotrasmettitori inibitori o eccitatori. La maggior parte di ciò che consente al nostro sistema nervoso di reagire adeguatamente al danno o alla lesione è la messa a punto o l'inibizione di una varietà di processi. La sovraeccitazione del sistema nervoso è considerata un problema in una serie di diversi disturbi. Ad esempio, l'iperattivazione di un recettore NMDA può anche essere correlata a disturbi affettivi, anomalie simpatiche e persino tolleranza agli oppiacei.
Anche il normale dolore nocicettivo, in una certa misura, attiva il recettore NMDA e si ritiene che porti al rilascio di glutammato. Tuttavia, nel dolore neuropatico, l'ipersensibilità al recettore NMDA è fondamentale.
Con altri tipi di dolore cronico, come la fibromialgia e il mal di testa di tipo tensivo, alcuni dei meccanismi attivi nel dolore infiammatorio e neuropatico possono anche creare anormalità simili nel sistema del dolore, tra cui sensibilizzazione centrale, maggiore eccitabilità delle vie somatosensoriali e riduzioni in meccanismi inibitori del sistema nervoso centrale.
Sensibilizzazione periferica
La cicloossigenasi (COX) svolge anche una funzione essenziale nelle sensibilizzazioni sia periferiche che centrali. COX-2 è uno degli enzimi indotti durante il processo infiammatorio; COX-2 converte l'acido arachidonico in prostaglandine, che aumenta la sensibilità dei terminali nocicettori periferici. Virtualmente, l'infiammazione periferica causa anche la produzione di COX-2 dal CNS. I segnali dei nocicettori periferici sono parzialmente responsabili di questa sovraregolazione, ma sembra anche esserci una componente umorale alla trasduzione dei segnali del dolore attraverso la barriera emato-encefalica.
Ad esempio, in modelli sperimentali, COX-2 viene generato dal SNC anche se gli animali ricevono un blocco nervoso sensoriale prima della stimolazione infiammatoria periferica. La COX-2 che viene espressa sui neuroni del midollo spinale del midollo spinale rilascia le prostaglandine, che agiscono sui terminali centrali, oi terminali presinaptici delle fibre sensoriali nocicettive, per aumentare il rilascio del trasmettitore. Inoltre, agiscono postsinapticamente sui neuroni del corno dorsale per produrre la depolarizzazione diretta. E infine, inibiscono l'attività del recettore della glicina, e questo è un trasmettitore inibitore. Pertanto, le prostaglandine creano un aumento dell'eccitabilità dei neuroni centrali.
Plasticità cerebrale e sensibilizzazione centrale
La sensibilizzazione centrale descrive i cambiamenti che avvengono nel cervello in risposta a ripetute stimolazioni nervose. Dopo ripetuti stimoli, quantità di ormoni e segnali elettrici cerebrali cambiano mentre i neuroni sviluppano un "ricordo" per reagire a quei segni. La stimolazione costante crea una memoria cerebrale più potente, quindi il cervello risponderà più rapidamente ed efficacemente quando si sottoporrà alla stessa stimolazione in futuro. Le conseguenti modifiche nel cablaggio e nella reazione del cervello sono indicate come plasticità neurale, che descrive la capacità del cervello di modificarsi prontamente o sensibilizzazione centrale. Pertanto, il cervello viene attivato o sensibilizzato da stimoli precedenti o ripetuti per diventare più eccitabile.
Le fluttuazioni della sensibilizzazione centrale avvengono dopo ripetuti incontri con il dolore. La ricerca sugli animali indica che l'esposizione ripetuta a una stimolazione dolorosa cambierà la soglia del dolore dell'animale e porterà a una risposta più forte al dolore. I ricercatori pensano che queste modifiche possano spiegare il dolore persistente che potrebbe verificarsi anche dopo un intervento chirurgico posteriore di successo. Anche se un disco erniato può essere rimosso da un nervo schiacciato, il dolore può continuare come un ricordo della compressione del nervo. I neonati sottoposti a circoncisione senza anestesia reagiranno in modo più profondo alla stimolazione dolorosa futura, come iniezioni di routine, vaccinazioni e altri processi dolorosi. Questi bambini hanno non solo una maggiore reazione emodinamica, nota come tachicardia e tachipnea, ma svilupperanno anche un pianto avanzato.
Questa memoria neurologica del dolore è stata studiata estensivamente. In una relazione sui suoi precedenti studi di ricerca, Woolf ha osservato che l'eccitabilità riflessa migliorata a seguito di danno o lesione del tessuto periferico non si basa su segnali di input periferici continui; piuttosto, ore dopo un trauma periferico, i campi recettivi del neurone del dorno spinale dorsale continuavano ad allargarsi. I ricercatori hanno anche documentato il significato del recettore NMDA spinale all'induzione e al mantenimento della sensibilizzazione centrale.
Significato per la gestione del dolore
Una volta stabilita la sensibilizzazione centrale, spesso vengono richieste dosi più elevate di analgesici per sopprimerla. L'analgesia preventiva o la terapia prima che il dolore progredisca, possono ridurre gli effetti di tutte queste stimolazioni sul SNC. Woolf ha dimostrato che la dose di morfina necessaria per interrompere l'ipereccitabilità centrale, somministrata prima di una breve stimolazione elettrica nociva nei ratti, era un decimo della dose necessaria per abolire l'attività dopo che era cresciuta. Questo si traduce in pratica clinica.
In uno studio clinico condotto su pazienti 60 sottoposti a isterectomia addominale, i soggetti che hanno ricevuto 10 mg di morfina per via endovenosa al momento dell'induzione dell'anestesia hanno richiesto una quantità significativamente minore di morfina per il controllo del dolore postoperatorio. Inoltre, la sensibilità al dolore intorno alla ferita, denominata iperalgesia secondaria, è stata ridotta anche nel gruppo pretrattato con la morfina. L'analgesia preventiva è stata utilizzata con un successo analogo in un assortimento di impostazioni chirurgiche, tra cui l'operazione prespinale e l'operazione postortopedica.
Una singola dose di 40 o 60 mg / kg di acetaminofene rettale ha un chiaro effetto di risparmio sulla morfina in chirurgia diurna nei bambini, se somministrata nell'induzione dell'anestesia. Inoltre, i bambini con analgesia sufficiente con paracetamolo hanno manifestato nausea e vomito postoperatori significativamente inferiori.
Gli antagonisti del recettore NMDA hanno somministrato analgesia postoperatoria quando somministrati prima dell'intervento. Esistono varie pubblicazioni in letteratura che supportano l'uso di ketamina e destrometorfano nel periodo preoperatorio. Nei pazienti sottoposti a ricostruzione del legamento crociato anteriore, il consumo di oppioidi analgesici controllati dal paziente 24-ora era significativamente inferiore nella categoria pre-operatoria del destrometorfano rispetto al gruppo placebo.
In studi di ricerca in doppio cieco, controllati con placebo, il gabapentin è stato indicato come analgesico premedicante per i pazienti sottoposti a mastectomia e isterectomia. Il gabapentin preoperatorio per via orale ha ridotto i punteggi del dolore e il consumo di analgesici postoperatorio senza gap negli effetti collaterali rispetto al placebo.
La somministrazione preoperatoria di farmaci anti-infiammatori non steroidei (FANS) ha dimostrato una diminuzione significativa dell'uso di oppiacei dopo l'intervento. I COX-2 sono preferibili a causa della relativa mancanza di effetti piastrinici e del significativo profilo di sicurezza gastrointestinale rispetto ai FANS convenzionali. Celecoxib, rofecoxib, valdecoxib e parecoxib, al di fuori degli Stati Uniti, somministrati prima dell'intervento riducono l'uso narcotico postoperatorio di oltre il 40 per cento, con molti pazienti che utilizzano meno della metà degli oppioidi rispetto al placebo.
Il blocco della conduzione nervosa nel periodo preoperatorio sembra impedire lo sviluppo della sensibilizzazione centrale. La sindrome dell'arto fantasma (PLS) è stata attribuita a un fenomeno di avvolgimento spinale. Pazienti con amputazione
spesso hanno dolore bruciante o formicolio nella parte del corpo rimossa. Una possibile causa è che le fibre nervose del moncone sono stimolate e il cervello interpreta i segnali come originari della porzione amputata. L'altro è il riarrangiamento all'interno delle aree corticali in modo tale che l'area dice che la mano ora risponde ai segnali provenienti da altre parti del corpo, ma continua a interpretarli come a venire per la mano amputata.
Tuttavia, per i pazienti sottoposti ad amputazione degli arti inferiori in anestesia epidurale, non uno dei pazienti 11 che hanno ricevuto il blocco epidurale lombare con bupivacaina e morfina per 72 ore prima dell'intervento ha sviluppato PLS. Per le persone sottoposte ad anestesia generale senza precedente blocco epidurale della lombare, 5 dei pazienti 14 ha avuto PLS alle settimane 6 e 3 ha continuato a sperimentare PLS nell'anno 1.
Woolf e Chong hanno notato che il perfetto trattamento preoperatorio, intraoperatorio e postoperatorio comprende "FANS per ridurre l'attivazione / centralizzazione dei nocicettori, anestetici locali per bloccare l'afflusso sensoriale e farmaci ad azione centrale come gli oppiacei." Diminuisce il dolore perioperatorio con tecniche preventive la soddisfazione, accelera lo scarico, risparmia l'uso di oppiacei, insieme a costipazione, sedazione, nausea e ritenzione urinaria, e può persino arrestare lo sviluppo di dolore cronico. Anestesisti e chirurghi dovrebbero prendere in considerazione l'integrazione di queste tecniche nelle loro pratiche quotidiane.
Quando il dolore si verifica a seguito di un danno o di una lesione in seguito a un intervento chirurgico, il midollo spinale può raggiungere uno stato ipereccitabile in cui si verificano reazioni eccessive di dolore che possono persistere per giorni, settimane o persino anni.
Perché la lesione localizzata risultante da un trauma provoca dolore cronico e intrattabile in alcuni pazienti? La lesione tissutale porta a una costellazione di variazioni dell'eccitabilità spinale, tra cui un innalzamento spontaneo elevato, una maggiore ampiezza e lunghezza della risposta, una soglia ridotta, una maggiore scarica a stimolazione ripetuta e campi espansivi recettivi. La persistenza di questi cambiamenti, che sono definiti collettivamente sensibilizzazione centrale, sembra essere fondamentale per il prolungato miglioramento della sensibilità al dolore che definisce il dolore cronico. Numerosi farmaci e / o farmaci, oltre al blocco neurale anestetico locale, possono limitare l'entità del windup del sistema nervoso centrale (SNC), come evidenziato dal calo del dolore e dal consumo diminuito di oppioidi nei modelli analgesici preventivi.
Insight di Dr. Alex Jimenez
La cura chiropratica è un'opzione di trattamento alternativo che utilizza regolazioni spinali e manipolazioni manuali per ripristinare in modo sicuro ed efficace e mantenere il corretto allineamento della colonna vertebrale. Studi di ricerca hanno determinato che i disallineamenti spinali, o sublussazioni, possono portare a dolore cronico. La cura chiropratica è comunemente utilizzata per la gestione del dolore, anche se i sintomi non sono associati a una lesione e / o condizione nel sistema muscolo-scheletrico e nervoso. Riorganizzando attentamente la colonna vertebrale, a chiropratico può aiutare a ridurre lo stress e la pressione dalle strutture che circondano il componente principale delle fondamenta del corpo esterno, fornendo in ultima analisi sollievo dal dolore.
Funzione e dolore del sistema nervoso enterico
Quando si tratta di ridurre l'uso di droghe e / o farmaci, compresi gli oppioidi, al fine di prevenire effetti collaterali come problemi di salute gastrointestinale, può essere in gioco la corretta funzione del sistema nervoso enterico.
Il sistema nervoso enterico (ENS) o sistema nervoso intrinseco è uno dei rami chiave del sistema nervoso autonomo (ANS) e consiste in un sistema nervoso a rete che modula il ruolo del tratto gastrointestinale. È in grado di agire indipendentemente dal sistema nervoso simpatico e parasimpatico, anche se potrebbe esserne influenzato. L'ENS può anche essere chiamato il secondo cervello.È derivato dalle cellule della cresta neurale.
Il sistema nervoso enterico nell'uomo è costituito da alcuni milioni di neuroni 500, inclusi i numerosi tipi di cellule di Dogiel, circa un duecentesimo della quantità di neuroni nel cervello. Il sistema nervoso enterico viene inserito nel rivestimento del sistema gastrointestinale, a partire dall'esofago e si estende fino all'ano. Le cellule di Dogiel, conosciute anche come cellule di Dogiel, si riferiscono a qualche tipo di tessuto surrenalico multipolare all'interno dei gangli simpatici prevertebrali.
L'ENS è in grado di svolgere funzioni autonome, come il coordinamento dei riflessi; anche se riceve una notevole innervazione nel sistema nervoso autonomo, funziona e può funzionare indipendentemente dal cervello e dal midollo spinale. Il sistema nervoso enterico è stato descritto come il "secondo cervello" per una serie di ragioni. Il sistema nervoso enterico può funzionare in modo autonomo. Normalmente comunica con il sistema nervoso centrale (SNC) tramite il parasimpatico, o tramite il nervo vago, e il simpatico, cioè attraverso i gangli prevertebrali, i sistemi nervosi. Tuttavia, studi sui vertebrati rivelano che quando il nervo vago viene reciso, il sistema nervoso enterico continua a funzionare.
Nei vertebrati, il sistema nervoso enterico include neuroni efferenti, neuroni afferenti e interneuroni, i quali rendono il sistema nervoso enterico capace di trasportare i riflessi e di agire come un centro di integrazione in assenza di input del CNS. I neuroni sensoriali riferiscono su condizioni meccaniche e chimiche. Il sistema nervoso enterico ha la capacità di cambiare la sua risposta sulla base di fattori quali la composizione nutriente e massiva. Inoltre, l'ENS contiene cellule di supporto simili all'astroglia del cervello e una barriera di diffusione attorno ai capillari che circondano i gangli, simile alla barriera emato-encefalica dei vasi sanguigni.
Il sistema nervoso enterico (ENS) gioca un ruolo fondamentale nei processi infiammatori e nocicettivi. Farmaci e / o farmaci che interagiscono con l'ENS hanno recentemente suscitato un notevole interesse per la loro capacità di regolare numerosi aspetti della fisiologia e fisiopatologia intestinale. In particolare, esperimenti su animali hanno dimostrato che i recettori attivati dalle proteine (PAR) possono essere essenziali per l'infiammazione neurogena nell'intestino. Inoltre, gli agonisti PAR2 sembrano indurre ipersensibilità intestinale e stati iperalgesici, suggerendo un ruolo per questo recettore nella percezione del dolore viscerale.
Inoltre, i PAR, insieme alle proteinasi che li attivano, rappresentano nuovi eccitanti bersagli per l'intervento terapeutico sull'ENS. Lo scopo delle nostre informazioni è limitato alla chiropratica e alle lesioni e condizioni spinali. Per discutere l'argomento, non esitate a chiedere al Dr. Jimenez o contattaci a 915-850-0900 .
A cura di Dr. Alex Jimenez
Argomenti aggiuntivi: Sciatica
Sciatica viene indicato come una raccolta di sintomi, piuttosto che una singola lesione e / o condizione. I sintomi del dolore del nervo sciatico, o della sciatica, possono variare in frequenza e intensità, tuttavia, è più comunemente descritto come un dolore improvviso, affilato (simile a coltello) o elettrico che si irradia dalla bassa schiena ai glutei, fianchi, cosce e gambe nel piede. Altri sintomi di sciatica possono includere, formicolio o bruciore sensazioni, intorpidimento e debolezza lungo la lunghezza del nervo sciatico. La sciatica colpisce più frequentemente individui di età compresa tra 30 e 50. Può spesso svilupparsi a causa della degenerazione della colonna vertebrale a causa dell'età, tuttavia, la compressione e l'irritazione del nervo sciatico causate da un rigonfiamento o Disco erniato, tra gli altri problemi di salute spinale, può anche causare dolore al nervo sciatico.
Lo strumento Find A Practitioner di IFM è la più grande rete di riferimento in Medicina Funzionale, creata per aiutare i pazienti a localizzare professionisti di Medicina Funzionale in qualsiasi parte del mondo. I professionisti certificati IFM sono elencati per primi nei risultati di ricerca, data la loro vasta formazione in Medicina Funzionale
Prenotazione online e appuntamenti 24 ore su 7, XNUMX giorni su XNUMX *

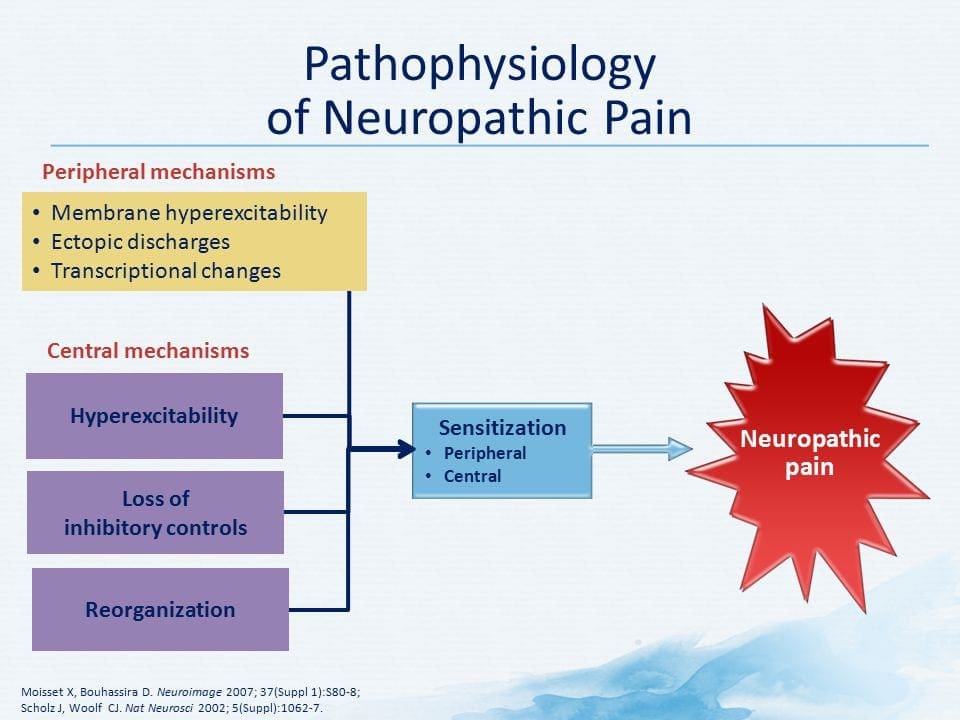
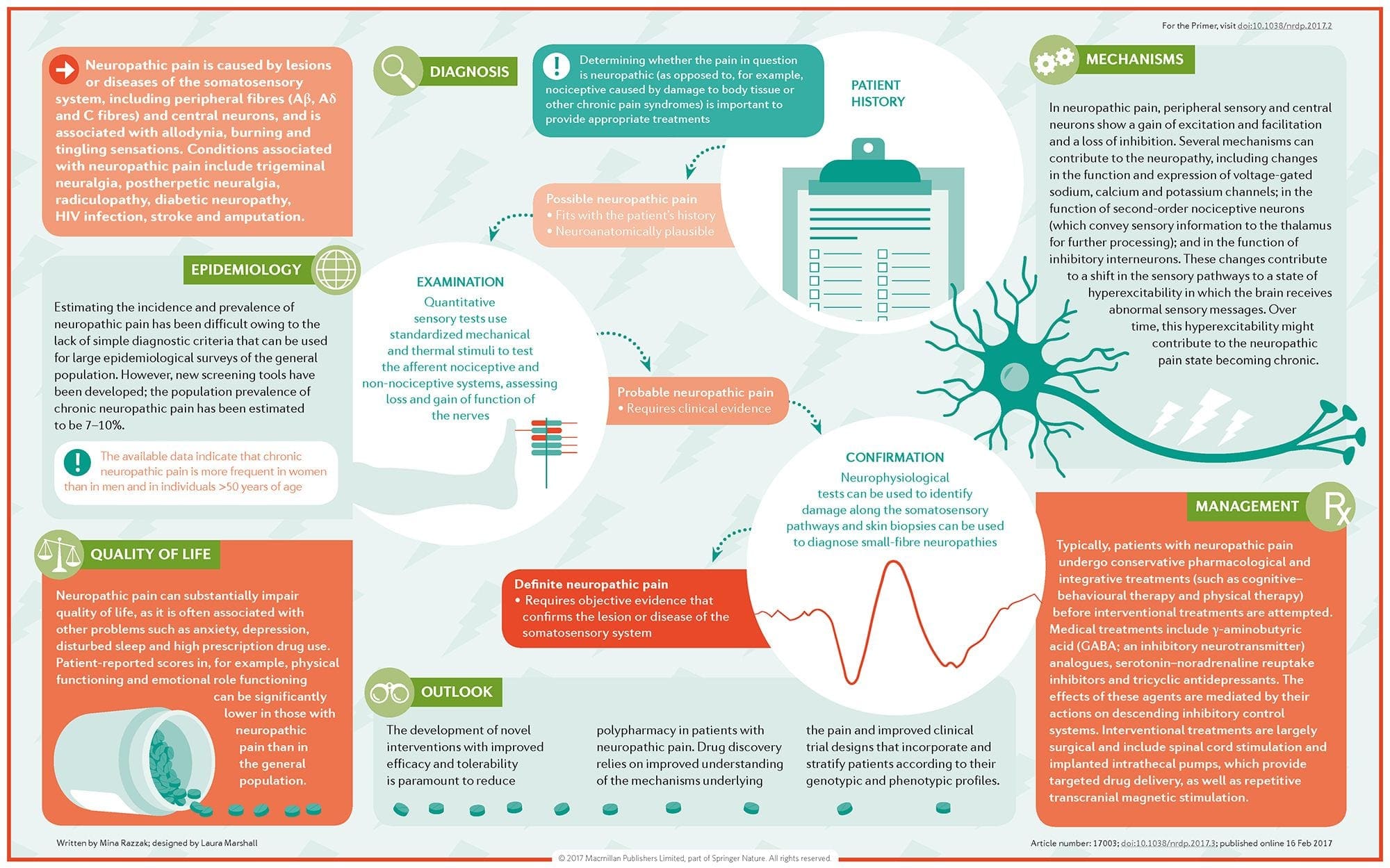




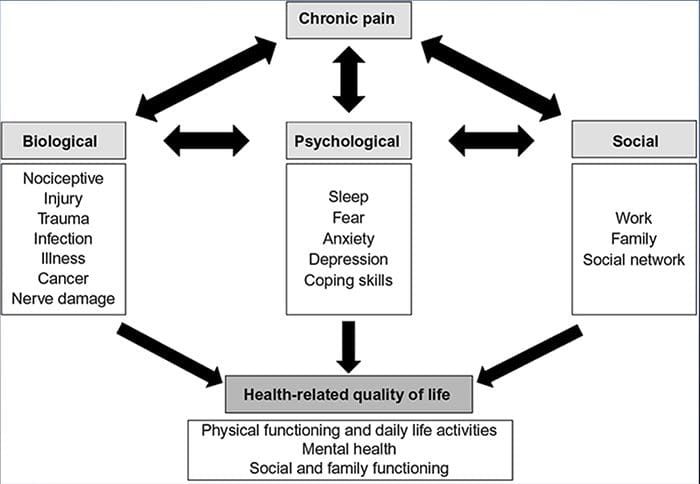
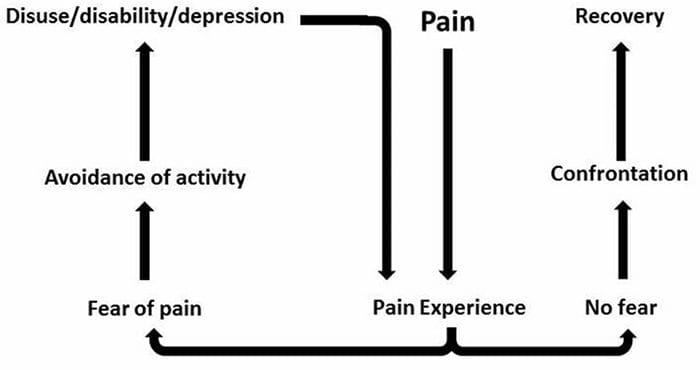
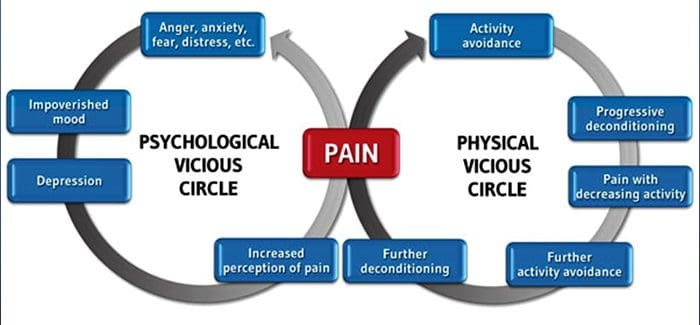
















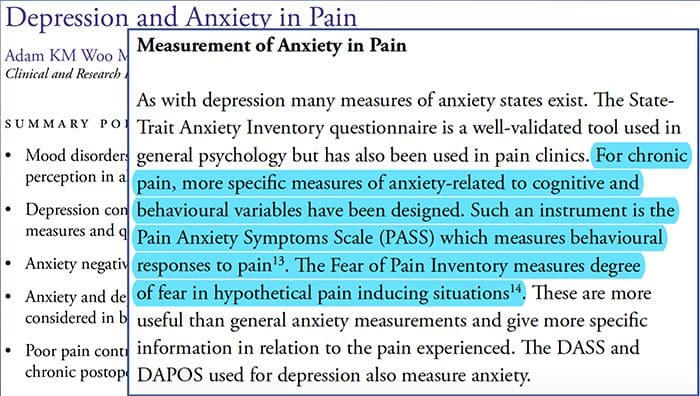


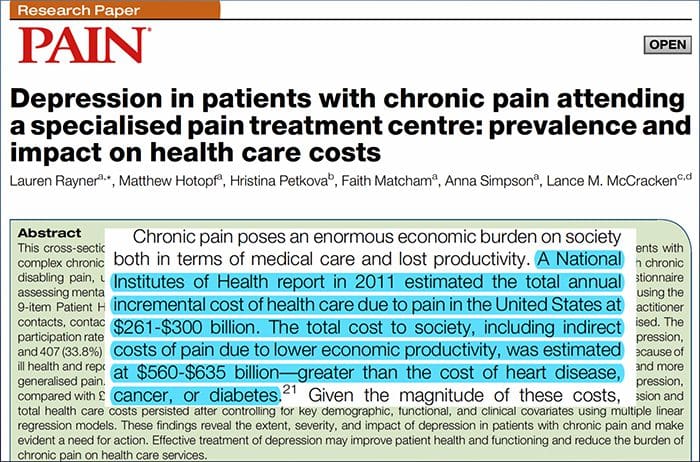



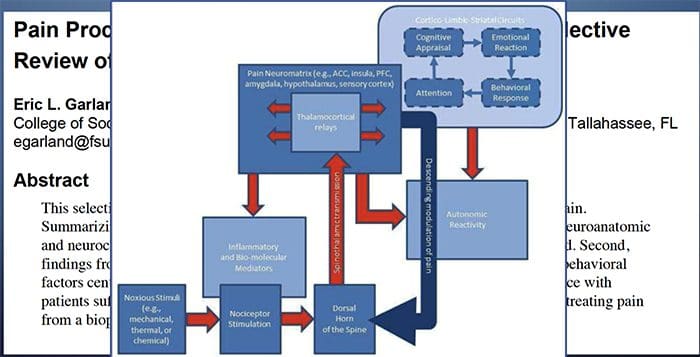






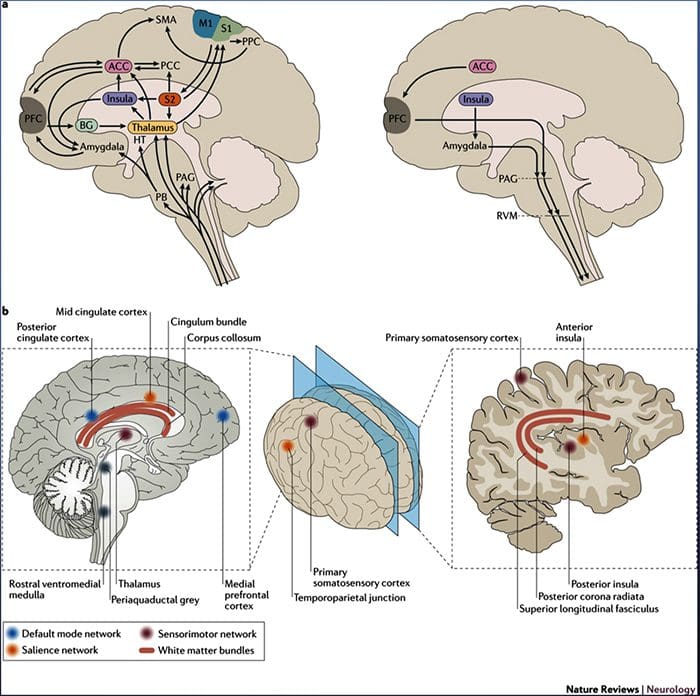

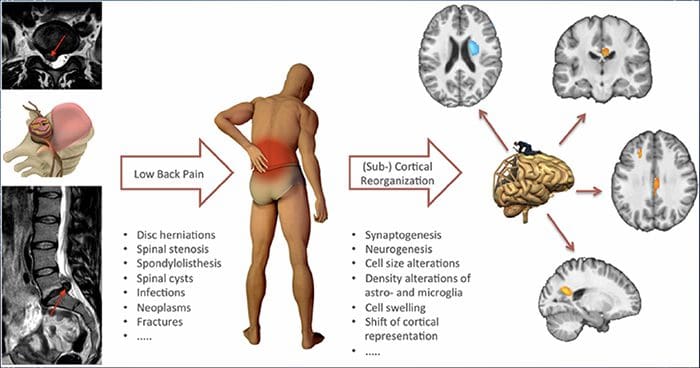

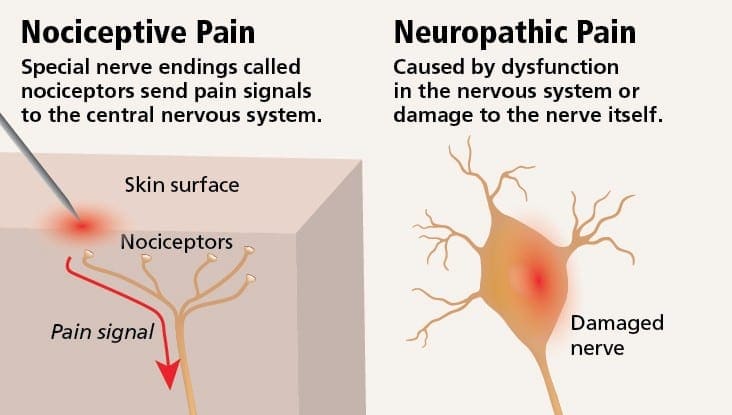
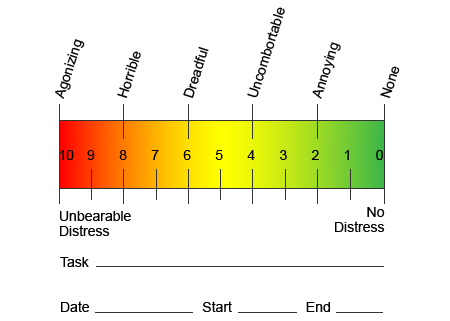


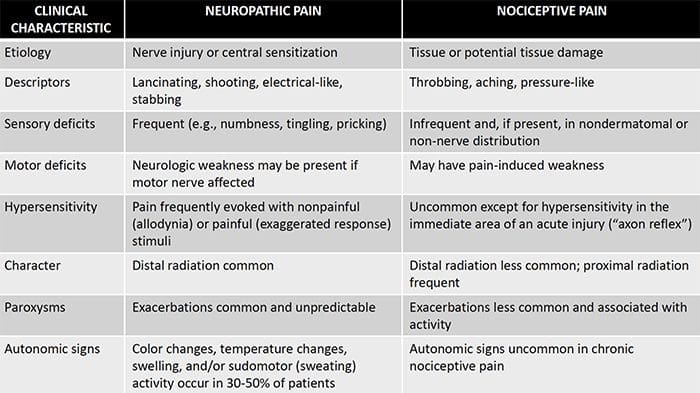
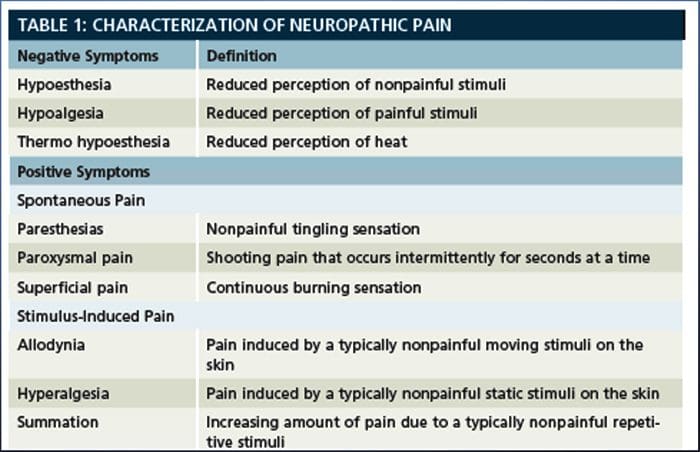
 PATOGENESI DEL DOLORE NEUROPATICO
PATOGENESI DEL DOLORE NEUROPATICO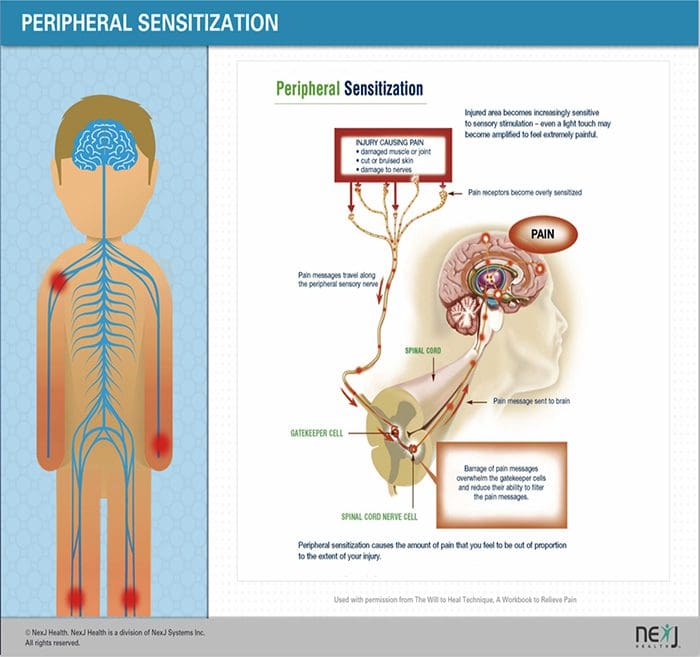
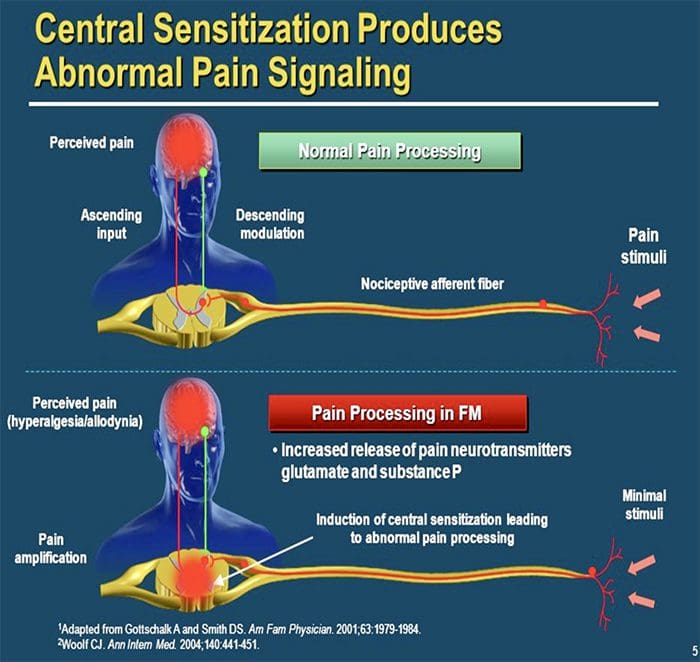
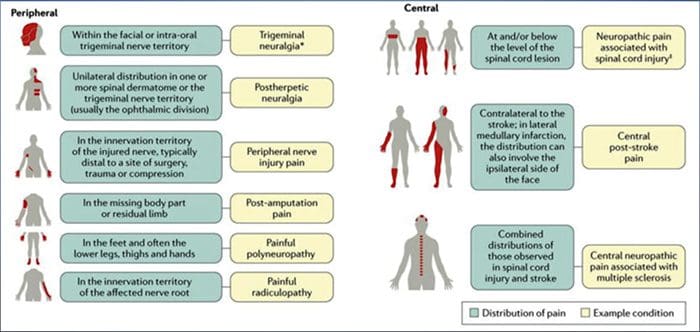 CAUSE COMUNI
CAUSE COMUNI
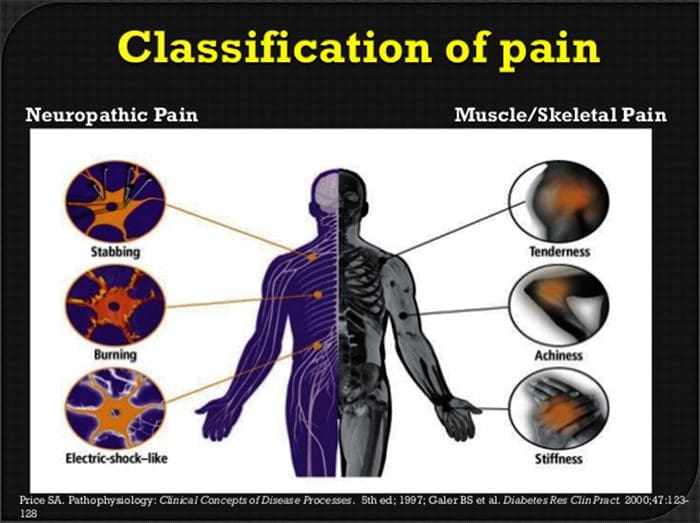
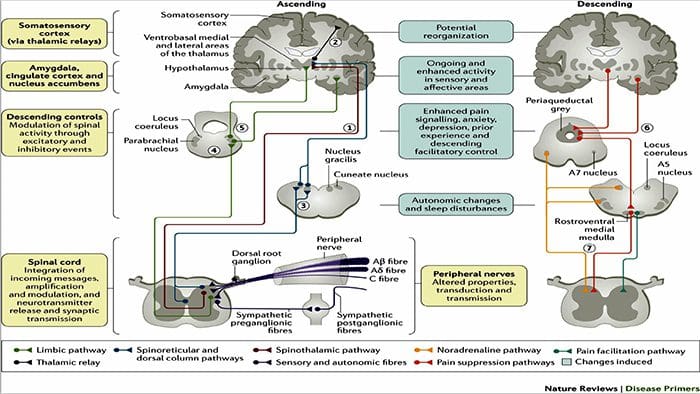
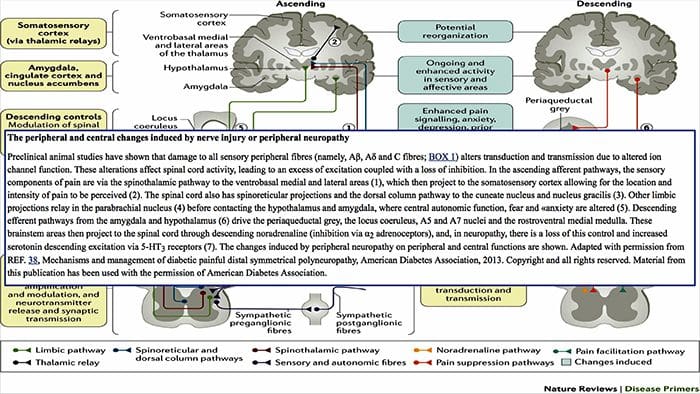

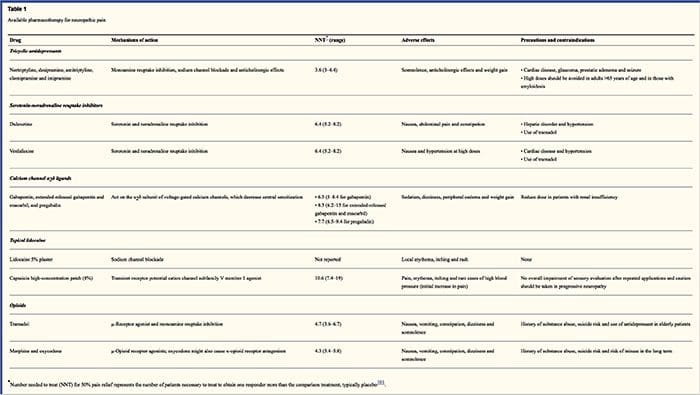




 DOLORE DEGLI ARTI FANTASMA E REALTÀ AUMENTATA
DOLORE DEGLI ARTI FANTASMA E REALTÀ AUMENTATA
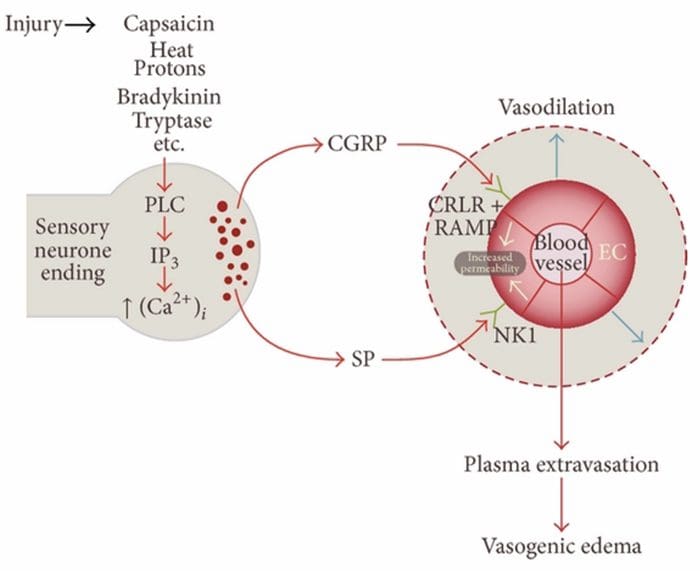 CAUSE COMUNI
CAUSE COMUNI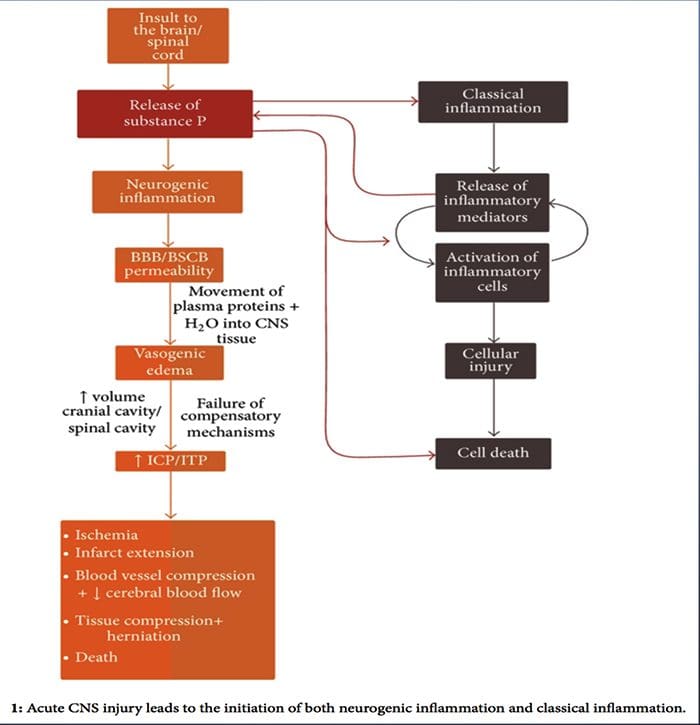


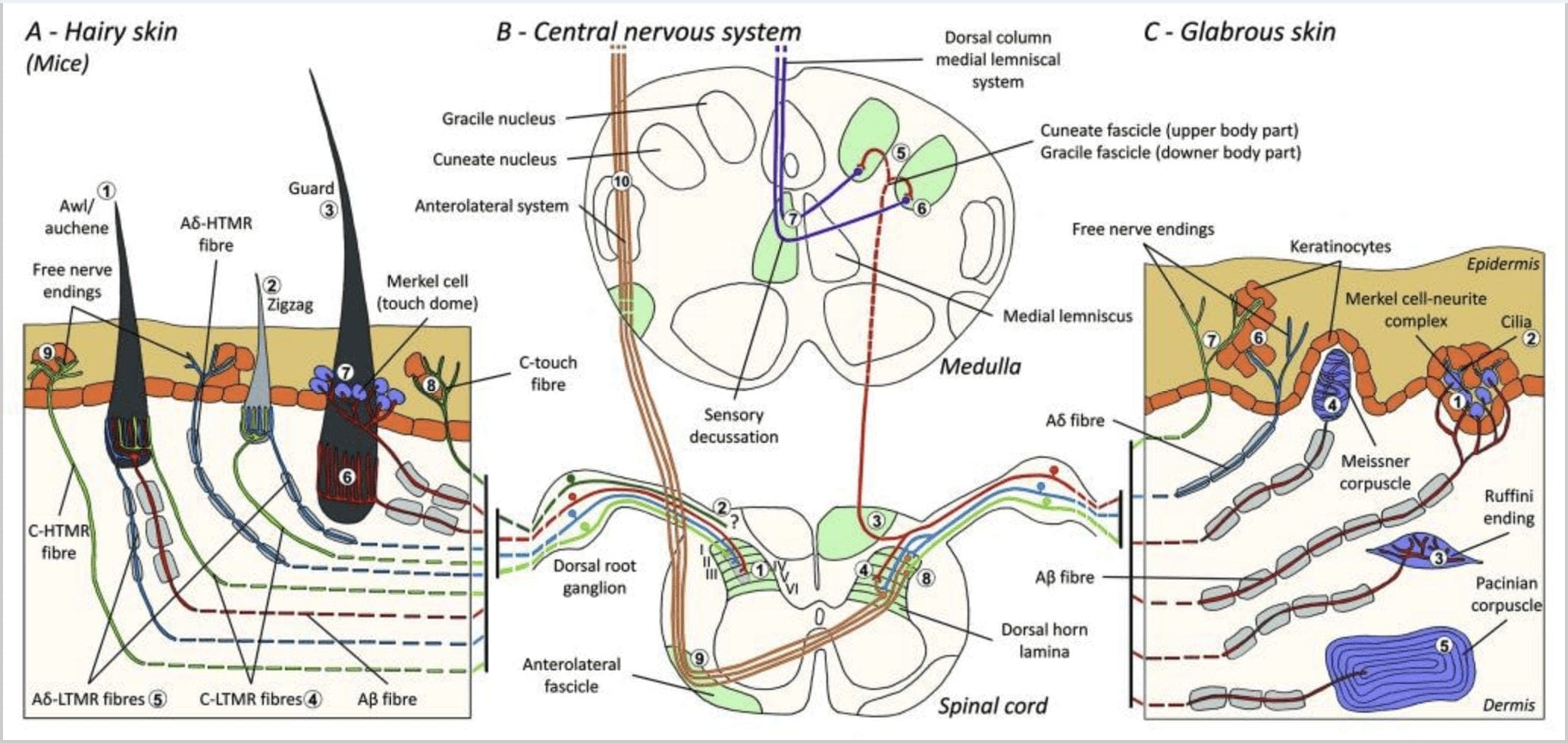
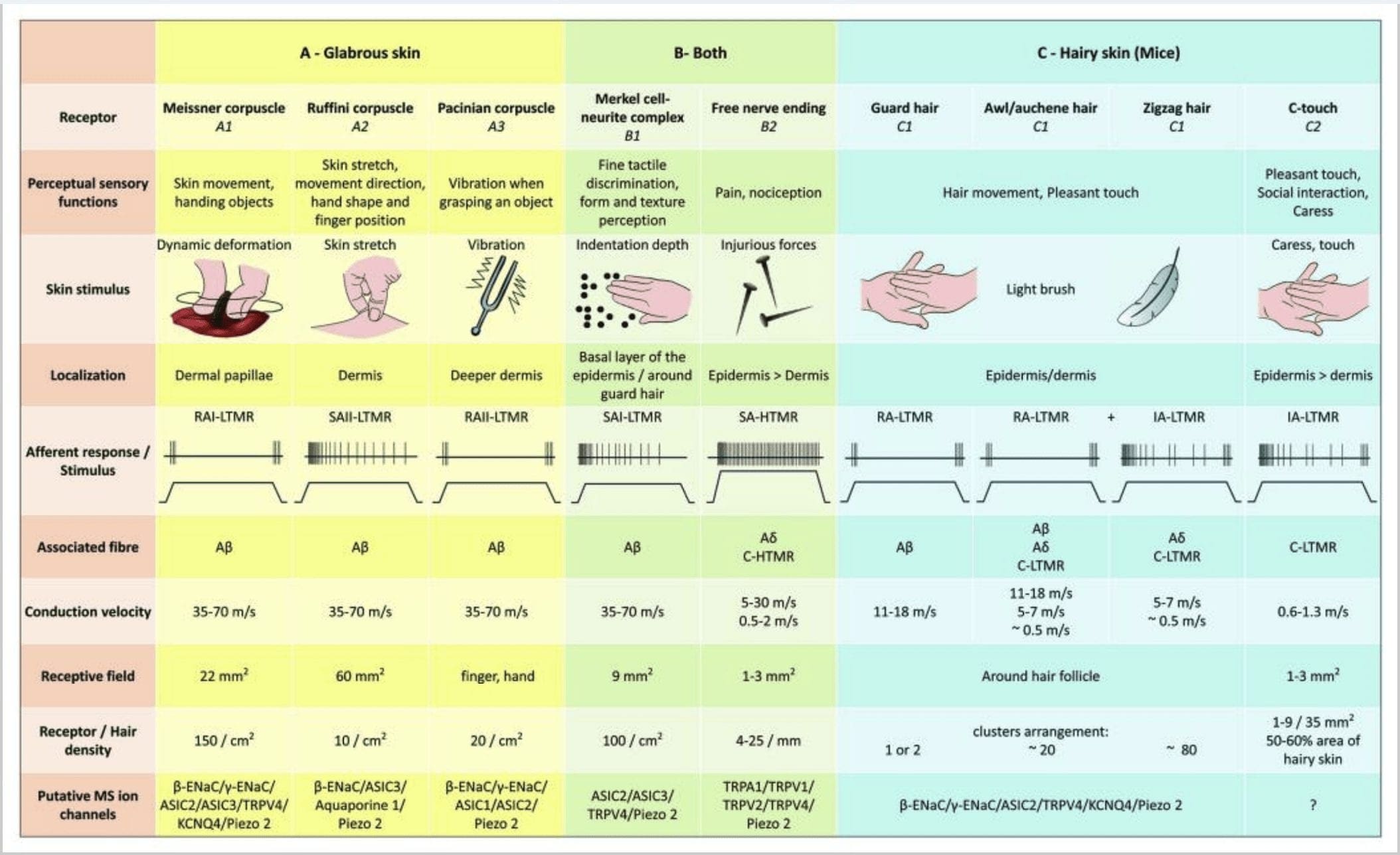

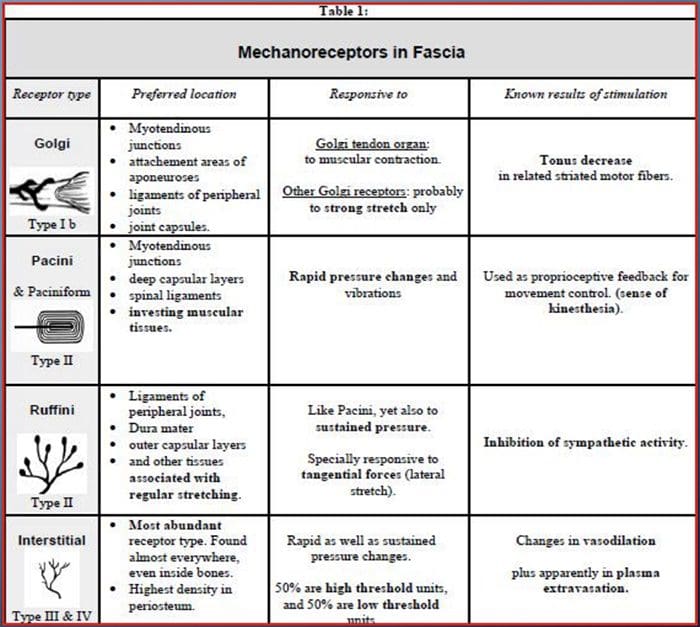 GATE CONTROL THEORY OF DOLORE
GATE CONTROL THEORY OF DOLORE
 PERCORSO LEMNISCALE DELLA MEDIA DELLA COLONNA DORSALE
PERCORSO LEMNISCALE DELLA MEDIA DELLA COLONNA DORSALE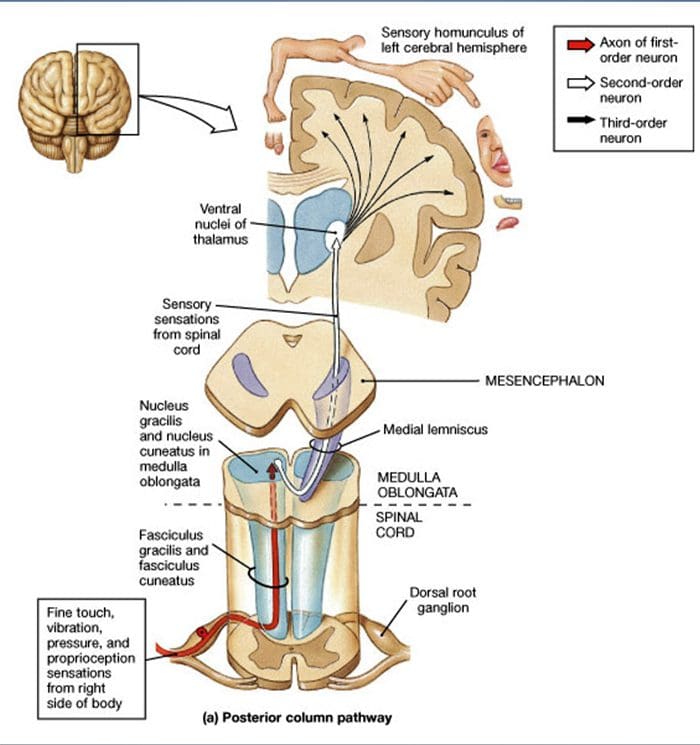 UTILIZZO DI MECCANORECETTORI PERIFERICI PER ALTERA PERCEZIONE DEL DOLORE
UTILIZZO DI MECCANORECETTORI PERIFERICI PER ALTERA PERCEZIONE DEL DOLORE
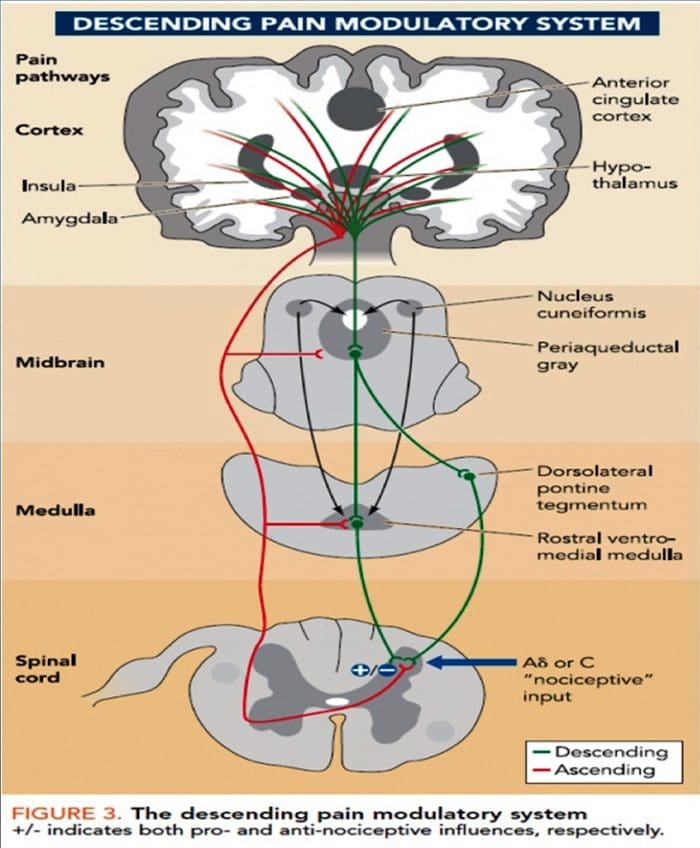
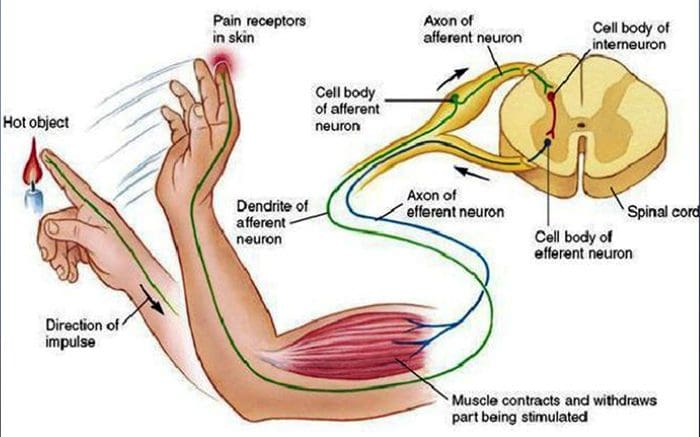
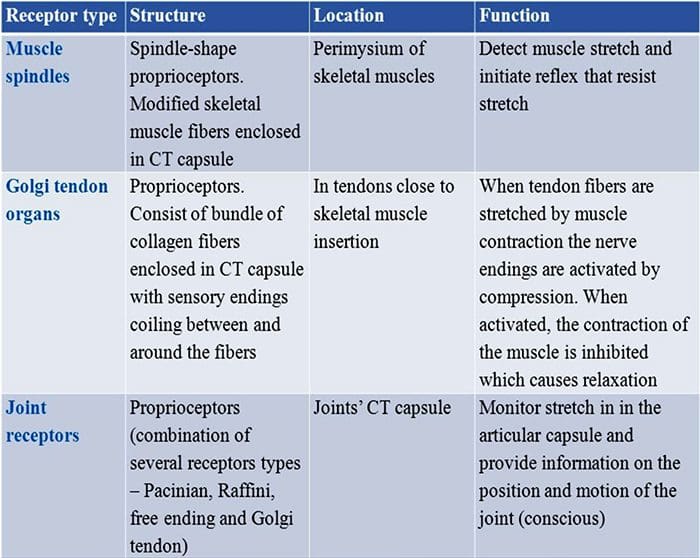
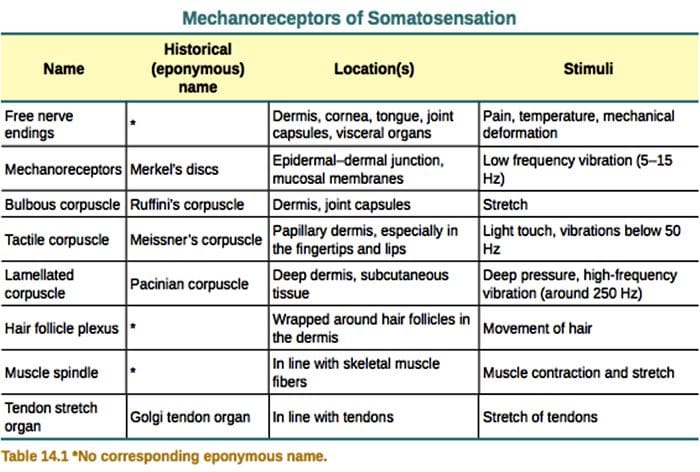 VIBRAZIONI
VIBRAZIONI
 TOCCO LEGGERO
TOCCO LEGGERO



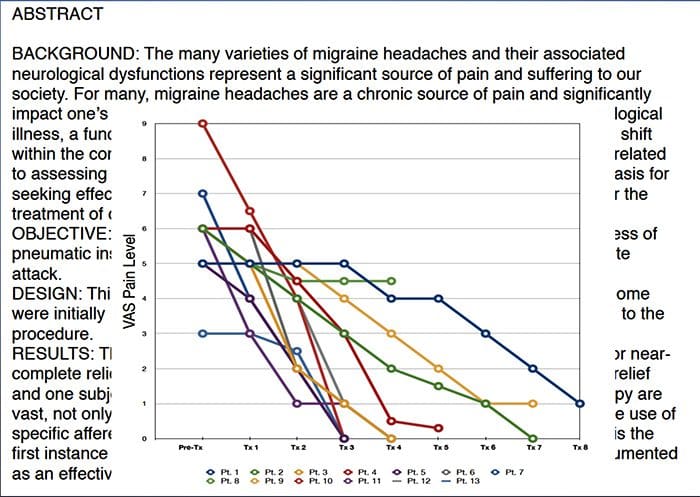


 CASE STUDY
CASE STUDY PUNTI SALIENTI DELL'ESAME DI FISICA
PUNTI SALIENTI DELL'ESAME DI FISICA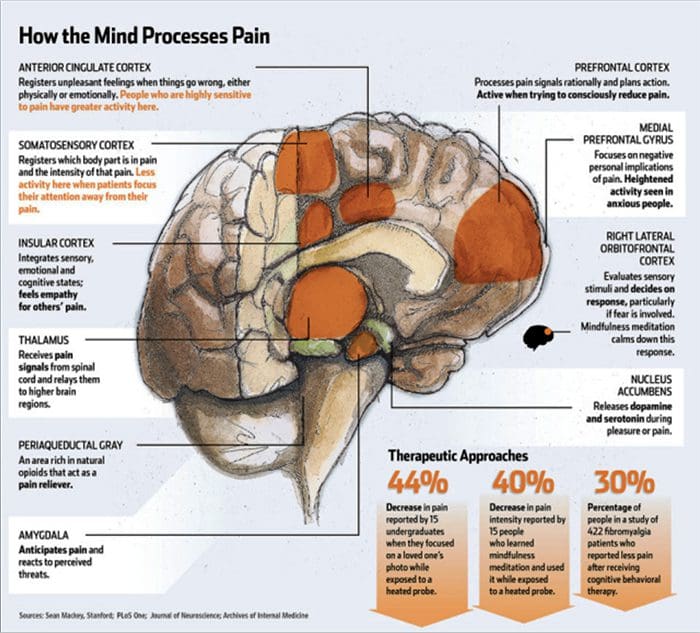 PUNTI SALIENTI DELL'ESAME DI FISICA
PUNTI SALIENTI DELL'ESAME DI FISICA
 DOPO DUE GIORNI
DOPO DUE GIORNI